
Le Transfert Horizontal de Gènes et les Gènes Orphelins – Créationnisme
Au fur et à mesure que les génomes de nouvelles espèces sont séquencés et ajoutés aux bases de données publiques, les chercheurs trouvent souvent des ensembles de gènes uniques, propres à certains types d’organismes. Par exemple, certains gènes sont présents uniquement dans certaines espèces, comme les rotifères1, les primates2, ou certaines plantes, sans qu’il n’existe d’équivalents dans d’autres lignées proches.
Cette spécificité génétique ne correspond pas aux attentes de la théorie de l’évolution qui postule un ancêtre commun et une divergence graduelle. On observe dans ces découvertes une difficulté croissante pour la théorie de l’évolution car elles mettent en lumière des différences non expliquées par des liens de parenté évolutifs clairs.
Le transfert horizontal de gènes (THG)
Le transfert horizontal de gènes (THG) est le processus par lequel un gène est transféré d’un organisme à un autre sans transmission héréditaire verticale (de parent à descendant). Si ce phénomène est largement observé chez les bactéries (qui échangent facilement des gènes pour par exemple développer une résistance aux antibiotiques), il est beaucoup plus controversé chez les organismes multicellulaires et complexes, comme les animaux ou les plantes.
Les évolutionnistes utilisent le THG de manière abusive pour expliquer l’apparition des gènes orphelins (gènes uniques sans homologues dans d’autres espèces) chez des organismes non apparentés, sans présenter de mécanismes clairs de transfert. C’est une manière d’argumenter la macro évolution que ne peuvent expliquer les autres mécanismes standard de l’évolution comme la sélection naturelle et les mutations génétiques.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
Nous apercevons cette tentative dans cette publication par exemple3:
Le THG a contribué à l’évolution de nombreux animaux, voire de tous, et le processus est en cours dans la plupart des lignées.
Ce discours est bien évidemment différent de ceux dont nous avions été assurés pendant des décennies sur les mutations4:
…Les mutations sont essentielles à l’évolution. Chaque caractéristique génétique dans chaque organisme fut initialement le résultat d’une mutation.
Le Dr Jeff Tomkins réagit à la déclaration (de Crisp, A. et al. 2015) sur le THG5 en affirmant:
Dans cette étude, les chercheurs n’ont pas réussi à prouver une quelconque partie de cette affirmation extravagante, ni à démontrer un quelconque mécanisme spécifique expliquant comment le THG aurait pu se produire. En fait, l’étude dans son ensemble a été gravement défectueuse sur plusieurs fronts clés.
Les gènes orphelins
Les gènes orphelins sont des éléments uniques et sans précédents clairs dans les lignées génétiques. Ils posent un problème pour la théorie de l’évolution car ils ne s’inscrivent pas dans le modèle classique d’une divergence graduelle à partir d’un ancêtre commun. Ces gènes, propres à certaines espèces, semblent « surgir » sans explication évolutive évidente, ce qui constitue un défi à l’explication évolutionniste. Ils sont à l’image des nombreux organismes dans le registre fossile qui apparaissent « soudainement » et « sans ancêtres » pour expliquer leur évolution supposée.
Pour expliquer leur origine, certains chercheurs avancent que ces gènes pourraient être issus de séquences d’ADN non codantes qui auraient acquis de nouvelles fonctions. D’autres, comme les auteurs de l’étude citée par Tomkins6, proposent qu’ils sont le produit du transfert horizontal de gènes (THG) — un processus par lequel des gènes sont transférés d’un organisme à un autre sans reproduction. Cependant cette explication est inadéquate car elle repose souvent sur des similarités partielles et n’explique pas comment ces gènes se seraient intégrés de manière fonctionnelle dans des réseaux biochimiques complexes.
Des gènes vitaux pour les organismes
Tomkins souligne que beaucoup de ces gènes sont impliqués dans des fonctions métaboliques essentielles ce qui est incompatible avec l’idée qu’ils soient des gènes « étrangers » provenant du THG. En effet, ces gènes codent pour des protéines enzymatiques cruciales et sont intégrés dans des réseaux biochimiques essentiels. Cela indique qu’ils sont conçus pour fonctionner de manière intégrée et ne sont pas des éléments ajoutés de manière aléatoire ou opportuniste. C’est pourquoi il est peu probable que des gènes essentiels aient pu être importés via le THG sans perturber la biologie de l’organisme hôte.
La plupart des cas de THG connus impliquent des relations hôte-parasite spécifiques qui facilitent le transfert de gènes, une situation absente dans de nombreux cas où l’on retrouve ces gènes orphelins.
Les gènes clés, notamment ceux impliqués dans le métabolisme, doivent être bien intégrés dans les processus cellulaires complexes pour fonctionner correctement. Un transfert aléatoire pourrait difficilement réussir une telle intégration sans perturber l’équilibre biochimique de l’organisme receveur.
Une méthode inadéquate pour identifier les gènes « étrangers »
Tomkins explique que les chercheurs n’ont pas utilisé la séquence génétique complète pour identifier les gènes « étrangers » (d’origine microbienne) mais se sont concentrés sur des similitudes partielles dans les régions protéiques codées par ces gènes. Cela signifie que la comparaison ne prend en compte que certains segments de protéines et laisse de côté de grandes portions de la séquence génétique, ce qui peut donner des résultats trompeurs. Cette méthode est inadéquate pour déterminer l’origine réelle des gènes car elle néglige la structure complète et les autres éléments critiques des gènes d’animaux.
Les gènes des mammifères sont particulièrement complexes. En moyenne, seuls 10 % de la séquence génétique d’un gène animal codent pour des protéines ; le reste est constitué de régions régulatrices sophistiquées qui influencent comment et quand le gène est exprimé, ainsi que la façon dont il interagit dans le réseau cellulaire. En revanche, les gènes microbiens manquent de cette complexité et de ces séquences régulatrices. En omettant ces différences, les chercheurs ont faussé leurs conclusions en ignorant que les gènes d’animaux et microbiens diffèrent non seulement par leur séquence mais aussi par leur fonction et structure générale.
En fait, ils ont admis que leur affirmation selon laquelle la provenance étrangère du gène est purement hypothétique, en déclarant que:
« la certitude absolue dans l’attribution de la plupart des THG est impossible à atteindre »
Le transfert horizontal de gène dans la nature
Dans la nature, le THG se produit presque exclusivement dans des contextes de relations étroites entre un hôte et un parasite, comme lorsqu’une bactérie parasite insère son ADN dans l’organisme hôte. Par exemple, la bactérie Wolbachia peut intégrer de grandes portions de son ADN dans les cellules reproductrices d’insectes ou de vers. Ces situations sont très spécifiques et nécessitent un contact biologique prolongé et intime, où le parasite peut entrer dans les cellules de l’hôte. Tomkins note qu’en dehors de telles relations biologiques précises, le THG ne se produit pas de manière naturelle et reste une exception chez les organismes complexes, tels que les animaux multicellulaires.
Pour que le matériel génétique étranger intégré dans un organisme soit transmis aux générations suivantes, il doit cibler les cellules germinales, c’est-à-dire les cellules qui produisent les gamètes (spermatozoïdes et ovules). Si un gène étranger n’atteint pas ces cellules, il ne pourra pas être hérité par la descendance. Cette contrainte impose un défi de taille pour expliquer comment des gènes « étrangers » auraient pu entrer et s’intégrer de manière stable dans des espèces complexes à travers des lignées entières.
Aucun mécanisme clair n’a été proposé pour expliquer comment des centaines de gènes auraient pu réussir ce processus de manière indépendante sans un lien hôte-parasite spécifique.
En plus du ciblage des cellules germinales, ces gènes transférés devraient être intégrés dans des réseaux biologiques existants de manière fonctionnelle, ce qui est très improbable sans un mécanisme bien défini. Le simple fait de transférer un gène ne garantit pas son bon fonctionnement dans le nouvel organisme, surtout si ce gène est essentiel au métabolisme ou à d’autres fonctions biologiques complexes.
Chez les eucaryotes complexes comme les mammifères, les barrières biologiques entre espèces rendent le THG moins courant et souvent non fonctionnel. Les bactéries doivent cibler des cellules spécifiques (comme les cellules germinales) pour que le transfert soit héréditaire, ce qui est rare et souvent spécifique aux organismes plus simples ou aux cellules immunitaires qui peuvent capter de l’ADN environnemental sans l’intégrer dans le génome de manière stable.
Arbre de la vie ou verger de la vie?
Dans l’article intitulé « Incomplete lineage sorting and other ‘rogue’ data fell the tree of life »7, Jeffrey Tomkins et Jerry Bergman examinent des éléments génétiques qui semblent contredire le modèle évolutionniste traditionnel de « l’arbre de la vie » tel que proposé par Darwin avec son fameux « I think« .
Problème de tri de lignées incomplètes (Incomplete Lineage Sorting – ILS)
L’ILS est un phénomène dans lequel les arbres phylogénétiques construits à partir de séquences ADN montrent des divergences et des discordances, même entre des espèces prétendument proches. Ces discordances, particulièrement fréquentes avec des données provenant de lignées non interfertiles et d’organismes éloignés, sont difficiles à concilier avec l’idée d’un ancêtre commun.
MicroARN et gènes restreints par taxon
Les microARN et les gènes dits « taxonomiquement restreints » apparaissent soudainement dans certains groupes biologiques sans précurseurs évolutifs. Ces éléments génétiques, souvent spécifiques à un groupe, génèrent des arbres phylogénétiques très différents de ceux basés sur les gènes codants standards, ajoutant un niveau de complexité qui défie la théorie de l’ascendance commune.
Données « aberrantes » et méthodes de lissage statistique
L’article critique l’utilisation de modèles mathématiques et statistiques pour « lisser » les données discordantes et produire des arbres phylogénétiques conformes au modèle évolutif. Ces techniques biaisent les analyses en masquant des données gênantes qui pourraient autrement contredire les hypothèses évolutionnistes.
Modèle du « Verger » créationniste
En opposition au modèle de l’arbre de la vie, Tomkins et Bergman, comme d’autres créationnistes, proposent l’idée d’un « verger de la création », où des « types créés » distincts auraient subi des variations horizontales sans nécessairement se transformer en de nouvelles espèces par évolution verticale. Ce modèle est présenté comme plus compatible avec les preuves de mosaïques d’ADN et des variations propres à certains groupes.
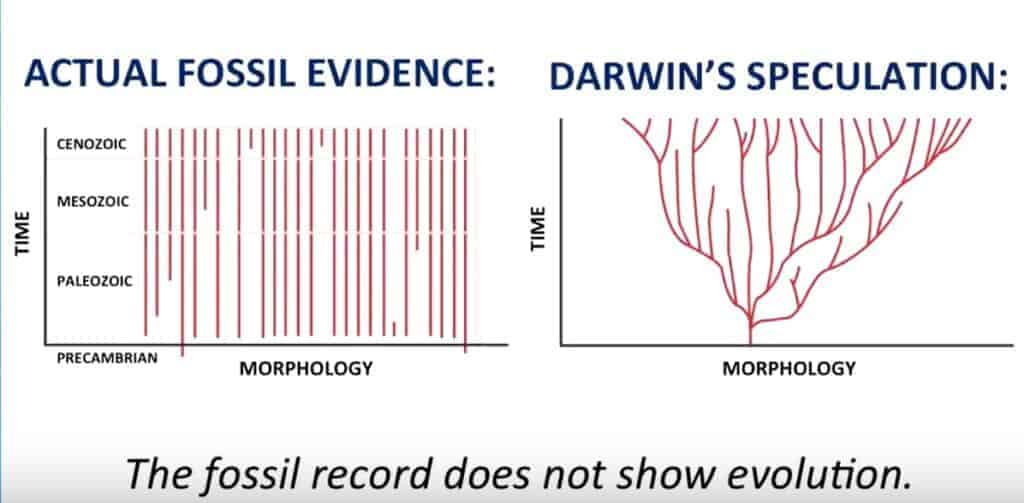
L’arbre de la vie et les « branches indépendantes »
L’un des problèmes avec l’arbre de la vie évolutionniste concerne les gènes orphelins qui ne semblent pas être reliés au tronc. Dans une étude de 2009 sur le sujet, Khalturin et al. ont noté que:
« les analyses comparatives du génome indiquent que chaque groupe taxonomique étudié jusqu’à présent contient 10 à 20 % de gènes qui n’ont pas d’homologues reconnaissables chez d’autres espèces ».8
Dans une autre étude, Tautz et Domazet-Lošo affirment que:
« chaque lignée évolutive abrite des gènes orphelins qui n’ont pas d’homologues dans d’autres lignées et dont l’origine évolutive n’est que mal comprise ».9
Tomkins déclare10:
Les TRG [gènes taxonomiquement restreints] apparaissent soudainement dans des génomes sans précurseurs évolutifs et leur présence est un mystère expliqué uniquement par des scénarios imaginatifs sans preuve concrète de l’existence d’un tel mécanisme. Les deux principales propositions hypothétiques pour les TRG sont la « synthèse de novo » et le « transfert horizontal de gènes » (également appelé transfert latéral de gènes).
Étant donné que les évolutionnistes tentent d’invoquer la duplication des gènes comme méthode d’origine de la plupart des gènes du génome, les TRG ne correspondent pas à ce modèle car ils n’ont aucune autre séquence dans le génome à partir de laquelle ils auraient pu être copiés – ils n’ont également, par définition, aucun paralogue (séquences similaires dans le même génome).
Le concept de manipulation ou de mélange aléatoire de l’ADN pour produire un gène fonctionnel complexe est un processus purement mythique, sans preuve. En fait, il défie les lois les plus élémentaires de la probabilité et de l’information, récemment décrites en détail par Meyer.11
Une autre idée évoquée est celle du transfert horizontal de gènes [THG], dans lequel les TRG [gènes orphelins] d’un taxon proviennent d’un transfert d’un autre taxon par des moyens non sexuels. Étant donné que, par définition, les TRG n’ont pas d’homologues dans d’autres taxons, l’organisme donneur serait inconnu. Chez les bactéries, le transfert horizontal de gènes se produit bien que sa fréquence n’ait pas encore été déterminée avec précision par une variété de mécanismes propres aux procaryotes. Alors que le transfert de gènes d’un microbe à un hôte a été documenté chez les eucaryotes, le transfert de gènes entre métazoaires12 manque d’un mécanisme prouvé viable.
Expérience avec de petits animaux aquatiques
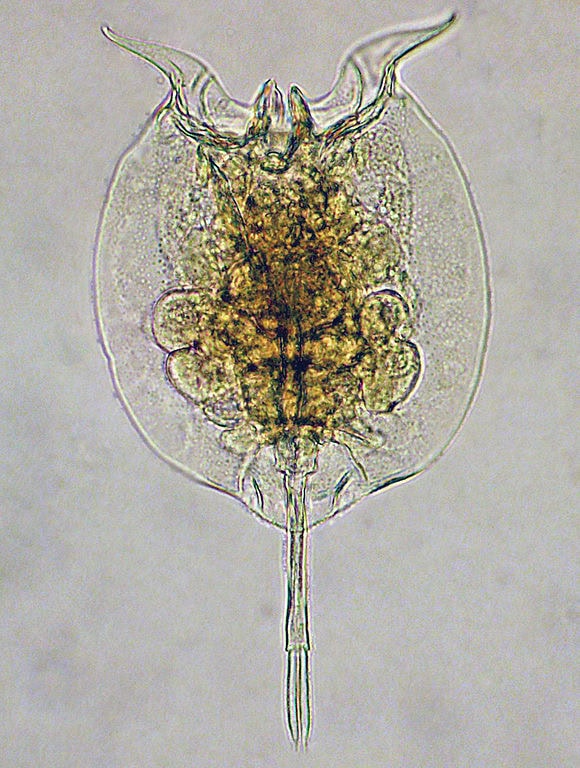
Dans une étude13, les chercheurs ont analysé plus de 61 000 gènes exprimés par les rotifères (de minuscules animaux aquatiques multicellulaires). Parmi eux, moins de la moitié présentaient des similitudes avec les séquences génétiques d’autres organismes tandis que le reste apparaissait comme propre et unique aux rotifères et difficile à expliquer par l’évolution. Le Dr Jeff Tomkins commente14:
Des 28 922 séquences pour lesquelles ils ont pu obtenir une correspondance dans une base de données publique de séquences d’ADN et de protéines d’autres créatures, une proportion significative (plus que chez toute autre créature séquencée) ne correspondait pas aux attentes évolutionnaires d’une descendance commune. Pour compliquer encore davantage ce tableau, les séquences de gènes de rotifères ont été trouvées dans un nombre varié de créatures non rotifères !
Les évolutionnistes ont deux options pour catégoriser ces correspondances génétiques inhabituelles en fonction de leurs présupposés naturalistes. D’abord, ils peuvent dire que ces gènes ont évolué indépendamment dans des créatures distinctes dans un processus hypothétique appelé « évolution convergente ». Cependant, dans les cas où des centaines de ces séquences d’ADN apparaissent dans plusieurs organismes, ce scénario devient si improbable que même les évolutionnistes ont beaucoup de mal à l’imaginer. La deuxième option est appelée « transfert horizontal de gènes », ou THG. Il s’agit du transfert de gènes, peut-être via une sorte de vecteur hôte microbien tel qu’une bactérie.
Le transfert horizontal de gènes (THG) est bien connu parmi les bactéries, où il est rendu possible grâce à des structures appelées pili. Ces pili sont des tubes protéiques qui permettent à une bactérie de se connecter à une autre et d’échanger de petits fragments d’ADN, un peu comme le partage de logiciels entre ordinateurs. Cette capacité d’échange génétique rapide est un avantage pour l’adaptation, comme dans le cas de la transmission de gènes de résistance aux antibiotiques.
Cependant, le THG entre une bactérie et un hôte multicellulaire est beaucoup plus rare et complexe. Il requiert un mécanisme permettant l’introduction et l’intégration stable d’un gène étranger dans le génome d’un organisme multicellulaire sans perturber son fonctionnement. Les cas observés sont souvent limités à des interactions symbiotiques ou parasitaires, dans lesquelles une bactérie passe du matériel génétique à un hôte spécifique avec lequel elle entretient une relation étroite.
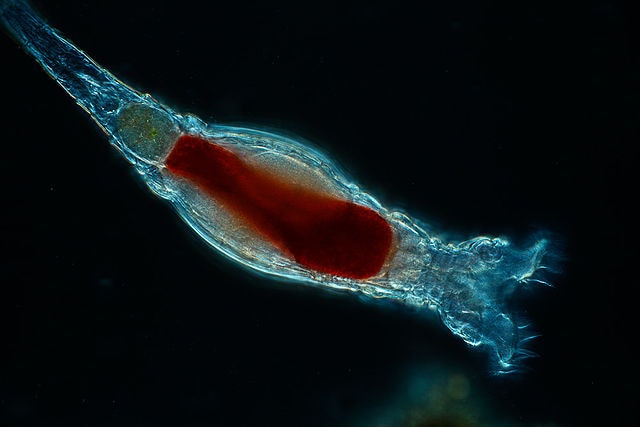
Dans le cas des rotifères(ces minuscules animaux aquatiques multicellulaires), Tomkins souligne la difficulté de justifier un transfert massif de centaines de gènes provenant de 533 sources génétiques différentes, dont les organismes n’ont pas de relations biologiques connues avec les rotifères. Pour que ce transfert massif de gènes ait eu lieu, il faudrait en théorie que chaque rotifère ait eu un lien hôte spécifique avec chacun de ces organismes – une hypothèse hautement improbable. Tomkins argumente ainsi que ce niveau de THG ne peut être expliqué de manière simple et pourrait plutôt indiquer une intégration intentionnelle de ces gènes dans le génome des rotifères.
Un autre aspect mis en avant est que les gènes « volés » supposément via le THG s’intègrent bien dans le système biochimique des rotifères ce qui suggère une adaptation spécifique et harmonieuse, difficile à expliquer simplement par le transfert de gènes aléatoires d’autres organismes. Tomkins note que le THG de gènes essentiels est théoriquement problématique, car ces gènes doivent s’intégrer dans un réseau biochimique complexe, ce qui rend improbable leur origine externe sans perturbations majeures. Il déclare:
Une étude distincte de 2012 a montré que les gènes natifs hautement exprimés ne pouvaient pas être partagés via THG, même entre bactéries, car ils perturberaient gravement la biochimie cellulaire essentielle. Et ce sont exactement les types de gènes qui ont été étudiés chez le rotifère.
…Il n’est pas connu non plus que le THG se produit en masse, et le THG des gènes essentiels est en théorie impossible.
L’obstacle majeur au transfert horizontal de gènes chez les organismes complexes
La publication citée par Tomkins est « high Expression Hampers Horizontal Gene Transfer (traduction: une expression élevée entrave le transfert horizontal de gènes) »15. Cet article explore un obstacle majeur au transfert horizontal de gènes (THG) : l’expression élevée des gènes. Les chercheurs ont étudié le phénomène dans le contexte des bactéries et des gènes qu’elles pourraient potentiellement partager avec d’autres organismes.
Les gènes qui sont fortement exprimés dans une cellule (c’est-à-dire que leur ARN et leurs protéines sont produits en grandes quantités) créent des contraintes lorsqu’ils sont transférés à un nouvel hôte. Ces gènes peuvent causer des déséquilibres biochimiques dans l’hôte récepteur, car leurs produits s’ajoutent aux réseaux métaboliques complexes déjà en place ce qui peut nuire au fonctionnement cellulaire.
Lorsque des gènes très exprimés sont introduits via le THG, leur intégration et leur expression dans le nouvel hôte nécessitent une compatibilité métabolique et des ajustements fins pour éviter une surcharge cellulaire. Les gènes faiblement exprimés ont plus de chances de réussir un transfert car leur intégration provoque moins de perturbations et de coûts énergétiques.
Cette contrainte peut limiter le THG pour certains types de gènes, en particulier ceux impliqués dans des fonctions essentielles et hautement régulées. Ce filtre naturel favorise le transfert de gènes moins exprimés, souvent impliqués dans des fonctions adaptatives ou accessoires, ce qui limite l’impact du THG sur les fonctions cellulaires de base et influence la façon dont les génomes évoluent et intègrent de nouvelles informations génétiques.
Symbiose et transfert de gènes chez les limaces de mer?
Dans un autre article du Journal of Creation16, Tomkins explore le cas fascinant des limaces de mer qui sont capables de photosynthèse, un phénomène assez rare et intrigant dans le règne animal. Les limaces de mer, notamment celles du genre Elysia, sont capables d’extraire des chloroplastes des algues dont elles se nourrissent et de les incorporer dans leurs propres cellules, leur permettant ainsi de produire de l’énergie à partir de la lumière solaire.

L’article soutient que cette capacité de photosynthèse remet en question certaines explications de l’évolution classique et de la théorie du THG, ce processus par lequel des gènes sont échangés entre organismes de différentes espèces, ce qui pourrait en théorie, expliquer comment les limaces de mer auraient pu acquérir cette compétence. Cependant, le niveau de complexité requis pour intégrer et maintenir les chloroplastes au sein des cellules des limaces ne s’explique pas simplement par des mécanismes évolutionnistes standards ou par le transfert de gènes. Cela pose la question de savoir si un tel niveau d’intégration n’indique pas une conception intelligente spécifique plutôt qu’un processus évolutif graduel.
L’exemple des limaces de mer photosynthétiques Elysia chlorotica (l’Élysie émeraude) nous permet de remettre légitimement en question le transfert horizontal de gènes (THG) comme explication pour les gènes communs que l’on retrouve chez des organismes sans liens évolutifs directs. Les scientifiques avaient initialement suggéré que le THG expliquait la présence de gènes d’algues chez ces limaces de mer, qui « volaient » des chloroplastes à leurs proies algales et les utilisent pour réaliser la photosynthèse.
De nouvelles recherches, qui ont séquencé l’ADN des cellules germinales de la limace via des œufs, n’ont trouvé aucun ADN d’algue. Cela signifie que les gènes des algues ne sont pas intégrés dans le génome héréditaire de la limace, réfutant ainsi l’hypothèse d’un THG chez E. chlorotica.
Les données originales soutenant le THG semblent maintenant s’expliquer par la présence d’eccADNs, de petites molécules d’ADN circulaires provenant du génome des algues et qui restent temporairement dans les cellules de la limace. Ces eccADNs pourraient jouer un rôle fonctionnel pour maintenir les chloroplastes algaux dans les cellules de la limace, mais sans être intégrés dans son génome.
Ce phénomène de photosynthèse chez E. chlorotica peut difficilement être expliqué par des processus évolutifs aléatoires mais suggère plutôt une conception intentionnelle et un système biologiquement complexe, un modèle de conception intelligente.
Conclusion
Le transfert horizontal de gènes, tout comme la duplication de gènes, la sélection naturelle, les mutations génétiques, la recombinaison génétique et autres mécanismes évolutifs, n’explique pas la macro évolution et l’innovation génétique. Il semblerait que les découvertes ne font qu’accentuer un modèle de conception intelligente en biologie. Il est évident que les futures découvertes et explorations des génomes continueront de nous enseigner les principes ingénieux et complexes qui sont mis en place pour permettre la vie. Tout cela doit nous (r)amener au Christ, créateur de toute cette diversité.
Mais souviens-toi de ton créateur pendant les jours de ta jeunesse, avant que les jours mauvais arrivent et que les années s’approchent où tu diras: Je n’y prends point de plaisir; (Ecclésiastes 12:1)
Notes et références:
- Les rotifères sont de minuscules animaux aquatiques multicellulaires, souvent microscopiques, qui vivent dans des milieux humides comme les étangs, les rivières et même les mousses et lichens. Ils mesurent généralement entre 0,1 et 0,5 mm et sont donc invisibles à l’œil nu. Leur corps est composé de plusieurs parties spécialisées, dont une « couronne » de cils autour de leur tête, qui ressemble à une roue et leur permet de se déplacer en tournant ou de faire circuler l’eau pour capter la nourriture. Cette caractéristique distinctive leur a valu le nom de « rotifères », du latin rota pour « roue » et ferre pour « porter ». Les rotifères se nourrissent de petites particules dans l’eau, comme des bactéries et des algues. Ils sont particulièrement fascinants pour les biologistes en raison de leur capacité à survivre dans des environnements extrêmes et même à supporter des périodes de dessiccation (séchage) en se mettant dans un état de dormance appelé anhydrobiose.
- Les primates sont un groupe d’animaux qui inclut les singes, les lémuriens, les gorilles et les chimpanzés (les évolutionnistes incluent les humains). Ils partagent des caractéristiques physiques et comportementales communes, comme des mains et des pieds qui permettent de saisir des objets, des pouces opposables (au moins pour certaines espèces), une vision développée pour voir en relief, et une intelligence plus avancée que celle de nombreux autres animaux.
- Crisp, A. et al. 2015. Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biology. 16: 50.
- Carlin, J.L. Mutations Are the Raw Materials of Evolution. Nature Education Knowledge 3 (10) : 10, 2011.
- https://www.icr.org/article/another-horizontal-gene-transfer-fairy.
- Crisp, A. et al. 2015. Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biology. 16: 50.
- https://creation.com/rogue-data-fell-tree-of-life.
- Khalturin, K., Hemmrich, G., Fraune, S., Augustin, R. and Bosch, T.C., More than just orphans: are taxonomically-restricted genes important in evolution?, Trends in Genetics 25(9):404-13, 2009 | DOI: 10.1016/j.tig.2009.07.006.
- Tautz, D. and Domazet-Loso, T., The evolutionary origin of orphan genes, Nature Reviews Genetics 12:692–702, 2011 | DOI: 10.1038/nrg3053.
- https://creation.com/rogue-data-fell-tree-of-life.
- Pour l’étude de Meyer: Meyer, S.C., Signature in the cell: DNA and the Evidence for Intelligent Design, 1st edn, HarperOne, 2009.
- Les métazoaires comprennent la grande majorité des animaux que nous connaissons, tels que les mammifères, les oiseaux, les reptiles, les poissons, les insectes, les crustacés, les étoiles de mer, les mollusques, etc..
- Boschetti, C. et al. 2012. Biochemical Diversification through Foreign Gene Expression in Bdelloid Rotifers. PLOS Genetics. 8 (11): e1003035.
- Insert your note here.
- Park, C. and J. Zhang. 2012. High Expression Hampers Horizontal Gene Transfer. Genome Biology and Evolution. 4 (4): 523-532.
- Solar-powered sea slugs defy evolution and horizontal gene transfer.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!