
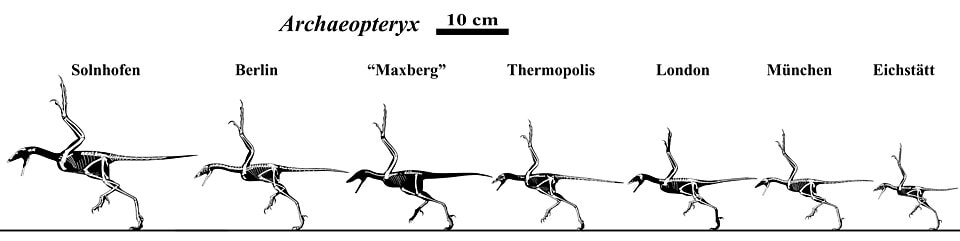
Archæoptéryx est-il l’ancêtre des oiseaux?
Archæoptéryx est un oiseau qui est daté à environ 150 Ma dans le référentiel évolutif (dans le Jurassique supérieur), il est traditionnellement présenté comme un « fossile transitionnel » ou un « intermédiaire structural » entre dinosaures et oiseaux par une majorité d’évolutionnistes. Dans cet article je me propose de réviser cet argument mais aussi plus globalement l’idée évolutive que certains théropodes auraient évolué en oiseaux.
- La prédiction darwinienne sur les fossiles de transition
- Des caractéristiques mosaïques
- Des dinosaures à plumes?
- Le vol optimisé d’Archæoptéryx
- Le système respiratoire
- Les dinosaures: des animaux à « sang froid »?
- Les cuisses internes des oiseaux
- L’acétabulum perforé
- Sinosauropteryx
- Confuciusornis
- Anchiornis
- Ichtyornis et Hesperornis
- Prudence sur les fossiles brandis comme des chaînons manquants
- Le plus ancien oiseau du registre fossile
- Des restes de protéines dans le fossile original d’Archaeopteryx
- Les traces d’oiseaux avant Archæoptéryx
- Des évolutionnistes dissidents?
- L’origine des dinosaures: un conundrum similaire comme avec l’origine des oiseaux?
- Conclusion
La prédiction darwinienne sur les fossiles de transition
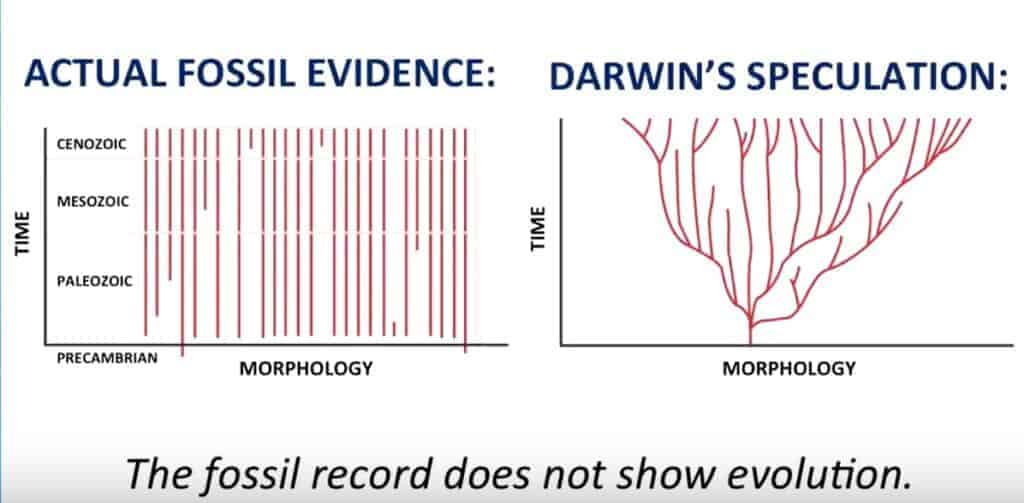
Dans « On the Origin of Species » (1859), Darwin écrivait que si sa théorie était vraie, le registre fossile devait contenir une profusion de formes transitionnelles, des chaînons nombreux montrant des gradations progressives. Par exemple, des fossiles à demi-ailes non fonctionnelles, ou à demi-poumons aviaires, représentant une progression morphologique continue, sans étape léthale.
L’idée est qu’il n’y avait pas d’organe suffisamment complexe pouvant empêcher une transition lente et graduelle. Cette idée est maintenant largement dépassée, tout (ou presque) est très complexe chez les êtres-vivants. Il faut beaucoup de choses immédiatement pour avoir une fonctionnalité de base. C’est un peu comme une voiture ou un avion, il faut des milliers, des millions de pièces pour avoir une fonction utile.
La nature « incrémentielle » de l’évolution est à ce titre problématique. Là où la théorie de Darwin postulait du temps long pour établir les êtres-vivants, on se rend compte qu’il faut aller très vite pour avoir tous les composants fondamentaux et indispensables à la fonctionnalité biologique.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
En fait, le problème de l’absence de fossiles transitionnels, ne se situe pas qu’au niveau des fossiles mais au niveau conceptuel, l’évolutonniste Stephen Jay Gould déclarait:
« L’absence de preuves (fossiles) pour les étapes intermédiaires entre les transitions majeures dans la conception organique, et en fait notre incapacité, même dans notre imagination, à construire des intermédiaires fonctionnels dans de nombreux cas, a été un problème tenace et persistant pour les récits graduels de l’évolution »
Gould, S.J., Is a new and general theory of evolution emerging? Paleobiology 6:119–130 (p.127), 1980.
Plus tard, il avait admis qu’il ne savait même pas à quoi pouvait ressembler les formes intermédiaires:
« Tous les paléontologues savent que les archives fossiles contiennent très peu de formes intermédiaires ; les transitions entre les grands groupes sont généralement abruptes.
Même si nous n’avons aucune preuve directe de transitions progressives, pouvons-nous inventer une séquence raisonnable de formes intermédiaires, c’est-à-dire d’organismes viables et fonctionnels, entre les ancêtres et les descendants ?
À quoi peuvent bien servir les stades initiaux imparfaits de structures utiles ?
À quoi sert une demi-mâchoire ou une demi-aile ?«
Gould, Stephen Jay. 1977. The Return of Hopeful Monsters. Natural History 86(6): 22–30.
Cette citation est marquante, puisqu’elle indique deux choses:
- Il n’y a « aucune preuve directe » de fossiles de transitions (pas d’observation)
- On ne peut même pas concevoir sur un papier ce qu’aurait pu être une transition entre ancêtres et descendants (pas de théorie non plus)
Il devait y avoir des transitions majeures nombreuses entre grands groupes, c’est à dire entre invertébrés et vertébrés, poissons et amphibiens, reptiles et oiseaux, etc. Ces transitions devaient être nombreuses et visibles dans les couches sédimentaires. Les couches plus anciennes devaient contenir les formes plus simples, et les fossiles de transition devaient combler les écarts entre les grands embranchements.
La formule de Gould « à quoi sert une demi-mâchoire ou une demi-aile ? » exprime le problème de la viabilité fonctionnelle, en effet, si une structure est incomplète, elle pourrait être un handicap plutôt qu’un avantage. Une « demi-aile » ou une « demi-mâchoire » n’est pas viable, parce que la fonction finale exige un ensemble complet et intégré. C’est pourtant ce qui était prédit par le darwinisme, on s’attendait à une évolution graduelle au fil des années avec complexification.
Cette tendance du simple au complexe n’a jamais été vérifiée dans le registre fossile. Par exemple il n’y a qu’à voir l’exemple des yeux des trilobites, « les yeux les plus complexes de tous les temps« , qui sont les …. « premiers yeux du registre fossile »… La complexité est donc là d’entrée de jeu.

Les invertébrés marins ne sont pas plus primitifs que les animaux qui apparaissent au dessus d’eux dans la colonne géologique, ils appartiennent à des biomes spécialisés et sont conçus de manière optimale pour remplir ces niches écologiques. Un singe (actuel ou fossile) n’est pas moins évolué qu’un humain, il est conçu pour une niche particulière (arboricole) et peut faire des choses que l’humain ne peut pas faire et inversement.
Il est totalement vain de raisonner en complexité quand les caractéristiques sont adaptées à une niche et à des fonctions particulières. Les microbes font de nombreuses choses essentielles pour les animaux et les humains, ils ne sont pas moins évolué, personne d’autres qu’eux ne peuvent faire ce qu’ils font.
Imaginez-vous voir clairement dans l’eau avec un champ quasiment de 360°. Certaines espèces (comme les Phacops) possédaient des yeux schizochroaux avec de grandes lentilles individuelles, leur donnant probablement une vision tridimensionnelle (stéréoscopique), en plus du large champ visuel.
Le même genre de constat est observé chez l’Archæoptéryx, qui pourrait être considéré, sous certains angles, comme ayant une palette plus riche que certains oiseaux dit « modernes ».
Dans le modèle créationniste la richesse génétique et la complexité étaient à leur paroxysme à la création. A la suite du péché d’Adam et Eve et du Déluge, les génomes se sont dégradés, la violence s’est accrue et l’environnement lui-même est devenu moins propice à la vie, cela a engendré des extinctions.

La majorité des trilobites étaient benthiques (vivant sur ou dans les sédiments au fond de la mer). C’est pourquoi ils font partie des premières créatures retrouvées, car le déluge de Noé a commencé dans les fonds océaniques avec l’éclatement de la croûte terrestre et le début de l’activité volcanique sous-marine, qui a provoqué l’ensevelissement de trilliards d’organismes invertébrés.
Des caractéristiques mosaïques

Les créationnistes considèrent Archaeopteryx comme un véritable oiseau, avec des plumes déjà parfaitement formées. Ses caractéristiques inhabituelles par rapport aux oiseaux observés « actuellement » (dents, griffes aux ailes, longue queue osseuse) sont des variations au sein des oiseaux. Nul besoin de les voir comme des traits de « transition ».
Les plumes de cet oiseau sont déjà aérodynamiquement optimisées, Archaeopteryx ne montre aucun processus de fabrication progressive. Il confirme l’observation récurrente dans le registre fossile « d’apparition soudaine ».
Le biais évolutif est à la base même du nom de l’oiseau, Archæoptéryx signifie « aile ancienne« , mais ses ailes n’ont rien d’archaïque.
La plupart des scientifiques l’ont accepté essentiellement comme un oiseau, jusqu’à récemment. Les scientifiques évolutionnistes placent désormais l’Archæoptéryx dans la catégorie imaginaire des dinosaures à plumes.1
Ses plumes ne constituent pas une transition entre les écailles et les plumes, car ses plumes sont abouties et optimisées. On n’y perçoit pas un stage intermédiaire. Certains évolutionnistes peuvent argumenter « l’évolution rapide », mais si on a que des écailles optimisées d’un côté et des plumes entièrement formées de l’autre, il est difficile de voir ce qu’il y a de transitionnel ou d’intermédiaire dans Archæoptéryx.
Une chauve-souris n’est pas une forme de transition entre l’oiseau et le mammifère terrestre, et un ornithorynque n’est pas une forme de transition entre le canard et le mammifère, même s’il présente certaines caractéristiques des deux.
Si donc les mosaïques de caractères de la chauve-souris et de l’ornithorynque ne nécessitent pas de relation évolutive, difficile de comprendre pourquoi la mosaïque d’Archæoptéryx devrait absolument refléter un intermédiaire entre reptile et oiseau.
John Morris, ancien président de l’Institute for Creation Research, écrivait dans son ouvrage de 2010, »
The Fossil Record*« :
« Il ne suffit pas qu’Archæoptéryx partage certaines caractéristiques squelettiques avec les reptiles, car c’est également le cas de tous les oiseaux et mammifères actuels. Cela implique-t-il pour autant une parenté? »
Tous les vertébrés terrestres partagent des traits hérités du plan tétrapode:
- vertèbres
- côtes
- mâchoire articulée
- doigts pentadactyles (ou dérivés)
Les chauves-souris, les baleines, les humains ont tous des traits « reptiliens » si on raisonne ainsi.
C’est une erreur évolutive fréquente: “X a des traits de Y, donc X descend de Y”. C’est un argument non sequitur.
L’évolutionniste Barbara Stahl déclarait:
« Puisqu’Archaeopteryx occupe une position isolée dans le registre fossile, il est impossible de dire si cet animal a donné naissance à des formes volantes plus avancées ou s’il ne représentait qu’une branche latérale par rapport à la lignée principale. »
Elle a ajouté dans une édition ultérieur (Dover 1985):
« La découverte de véritables fossiles d’oiseaux datant du Crétacé inférieur n’a fait que renforcer l’argument selon lequel le célèbre Archaeopteryx à plumes pourrait être une branche archaïque dérivée de la lignée avienne ancestrale. »
Stahl, B.J., p. Vertebrate History: Problems in Evolution (McGraw–Hill 1973).
Son idée est qu’Archæoptéryx était une branche latérale, et non directement une branche centrale ayant mené vers les oiseaux. Dans cette vue il n’est techniquement pas l’ancêtre mais un cousin. C’est la vue de Barbara Stahl. Cela affaiblit d’entrée de jeu la notion de « transition ».
Ancêtre hypothétique inconnu
|
|
Lignée principale
|
Oiseaux modernes
|
Branche latérale
|
Archaeopteryx
Si Archaeopteryx n’est pas un ancêtre direct, il ne démontre pas la transformation dinosaure vers oiseau. Il n’est qu’un exemple de mosaïque de traits (plumes + dents + queue osseuse), mais cela ne prouve pas qu’il ait donné naissance aux oiseaux modernes.
En fait, la notion de cousin elle-même est contestable. Ce qu’on a réellement c’est « deux lignes parallèles » mais aucun trait qui les unis par une relation quelconque. C’est la logique de la « forêt de la vie » créationniste VS « l’arbre de la vie » évolutionniste.
Il y a des oiseaux actuels comme le hoatzin et le touraco (dont les poussins ont des griffes aux ailes) ou des oiseaux fossiles du Crétacé comme Ichthyornis (qui avait des dents et qui arrive bien après Confuciornis qui avait un bec non denté) pour dire que la présence de dents et de griffes n’est pas un trait exclusivement reptilien. Les griffes d’oiseau ne viennent pas nécessairement des reptiles.
Il existe dans le registre fossile quelques espèces, comme Ichthyornis et Hesperornis, qui avaient un bec et des dents. Les oiseaux « modernes » ont perdu ou n’ont jamais eu de dents.
Archæoptéryx a une mosaïque de caractères, comme l’ornithorynque avec son bec de canard (d’oiseau), son oviparité (de reptile) et son lait et ses poils (de mammifères).

Ces trois groupes d’oiseaux cités n’ont pas nécessairement de lien évolutif entre eux, ils peuvent très bien s’expliquer par le « concepteur en commun » qui réutilise des modules dans diverses conceptions, cela évite de faire appel à ces multitudes de cas « d’évolution convergente » qui stipulent que des mutations aléatoires ont plusieurs fois produit la même chose.
Il n’y a pas de preuve que la mâchoire dentée d’Archæoptéryx ait été transformée en bec kératinisé, ce n’est pas nécessité par les données observées. L’idée d’une mâchoire ou d’un bec qui se serait formé progressivement reste spéculative car aucun fossile ne documente une transformation progressive.
Si on souhaite relier A et Z, il faut commencer par démontrer B, C, D et ainsi de suite. Si on a que l’un ou l’autre, on ne peut pas prouver qu’il y a eu une transformation, ils ont très bien pu être créés séparément.
Des dinosaures à plumes?
L’étude de Chen et al. (1998) décrit un fossile exceptionnel de la formation de Yixian en Chine qui est interprété comme un dinosaure à plumes et est présenté comme une preuve clé des liens évolutifs entre dinosaures théropodes et oiseaux.
Les plumes fossilisées peuvent être des artefacts (fibres de collagène, structures tégumentaires non aviennes) mais même si c’étaient des plumes (ce que le créationnisme n’interdit pas), cela montrerait seulement de la mosaïque de caractères et pas une transformation.
Des paléontologues séculiers ont identifié des structures filiformes faisant saillie sur le corps de certains fossiles de dinosaures. Ils affirment qu’il s’agit de « proto-plumes », précurseurs des véritables plumes. Alan Feduccia et ses collègues n’ont trouvé aucune preuve, dans les découvertes publiées en provenance de Chine, que ces structures filiformes soient des plumes, ni même des proto-plumes.
Ils ont déterminé que les prétendues proto-plumes étaient en réalité les restes de fines fibres de collagène provenant d’une peau partiellement décomposée. Leurs recherches ont notamment porté sur l’analyse de fibres de collagène cutanées en décomposition chez des reptiles, des requins et des dauphins modernes, ainsi que sur la comparaison de ces fibres avec celles de plusieurs dinosaures.2
Les plumes ne sont pas issues des écailles:
« Les preuves provenant de la biologie du développement portent un sérieux coup à la vision classique selon laquelle les plumes auraient évolué à partir d’écailles allongées. »
— Thain & Hickman, Dictionary of Biology, 2004, p. 265-266.
Une étude de 2017 parue dans la revue Biology Letters a anéanti le rêve du « poulet tyran »: T. rex était couvert d’écailles. Les nouvelles recherches:
“montrent sans aucun doute que T. rex avait une peau écailleuse”.
— Phil Bell, paléontologue (auteur de l’étude: Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution ), cité par B. Guarino, The Washington Post, 6 juin 2017.
Les peaux de dinosaures retrouvées sont radicalement différentes des plumes d’oiseaux.
« Un voile de doute a été jeté sur la théorie largement admise selon laquelle les dinosaures avaient des plumes et qu’ils ont donné naissance aux oiseaux modernes. »
— Lovgren, S., 2007, National Geographic News
Il y a également le cas des supposées plumes sur les avant-bras de Velociraptor ou d’autres théropodes. Cela fait référence à de petites bosses osseuses sur l’ulna (un os de l’avant-bras) qui sont interprétées comme des points d’attache pour les plumes de l’aile.
Les tubercules de plumes (connues sous le nom de « Quill Knobs » en anglais) sont de petites protubérances osseuses alignées le long de l’ulna. Chez les oiseaux modernes, elles servent de points d’ancrage aux ligaments qui fixent les plumes de vol (rémiges secondaires).
En 2007, des paléontologues ont décrit ces structures sur un fossile de Velociraptor. Les chercheurs ont observé 6 bosses alignées sur l’ulna, « exactement » à l’endroit où se fixent les plumes chez les oiseaux. Cette découverte a été publiée par l’équipe de Alan Turner. Selon les évolutionnistes cela indiquerait que Velociraptor possédait « probablement » de grandes plumes sur les bras. D’autres recherches ont par la suite été influencées et sont allées dans le même sens (le dakotaraptor par exemple).
Mais attention à aller trop vite en besogne. A savoir par exemple que les avant-bras d’autres Vélociraptors sont parfaitement lisses, il n’y a pas de protubérances. Si tous les membres de ce groupe possédaient des plumes nécessitant un ancrage aussi solide, il est étonnant que les spécimens les mieux conservés ne présentent pas ces protubérances. En réalité ces « protubérances » sont minuscules et lisses, il fallait les « grossir » pour les voir (en comparaison des protubérances de la plupart des oiseaux actuels).
Ce serait également une caractéristique étonnante, voire inutile sur un théropode. Chez l’oiseau, la rigueur et l’exigence du vol propulsé nécessite des attaches renforcées. Compte tenu de la petite taille des bras des droméosauridés, ils ne pouvaient pas servir pour le vol. A quoi pouvait servir de fixer fermement des plumes si le vol était impossible derrière? On a d’ailleurs pas trouvé de plumes à leurs côtés.
En réalité ces « protubérances » peuvent avoir d’autres fonctions que d’ancrer des plumes, par exemple servir de points d’attache pour les tissus conjonctifs. Le paléontologue évolutionniste Darren Naish (au sujet d’un Allosauridae) a déclaré:
« Les animaux présentent parfois des tubercules étranges, irrégulièrement espacés, disposés en lignes sur différents os, généralement situés le long des lignes intermusculaires (ils représentent probablement des sites d’attache partiellement ossifiés pour des feuillets tendineux ou des structures similaires). J’en ai observé sur des os de mammifères et sur un tibia de théropode. »
Concavenator: an incredible allosauroid with a weird sail (or hump)… and proto-feathers? Publié le 9 septembre 2010 – Darren Naish
Si les minuscules bosses sur les mammifères ne représentent pas nécessairement des plumes, alors l’interprétation des plumes n’est pas non plus obligatoire chez les dinosaures.
Joel Tay déclare:
« Malheureusement, comme nous l’avons souvent souligné, les présuppositions ont tendance à influencer l’interprétation des preuves. Dans ce cas précis, le désir de démontrer que les dinosaures ont évolué en oiseaux a conduit de nombreux évolutionnistes à interpréter de minuscules protubérances sur les os de dinosaures comme des tubercules de fixation des plumes. »
Il dit encore:
« Par exemple, chez Concavenator, les protubérances semblent se situer sur la face latérale du cubitus (vers l’extérieur du bras), plutôt qu’à l’arrière du bras comme on pourrait s’y attendre si ces tubercules servaient de points d’attache aux plumes de vol. Les véritables tubercules des plumes devraient également être régulièrement espacés, mais ces minuscules protubérances sont souvent irrégulièrement réparties chez ces dinosaures. »
Brian Thomas et Jonathan Sarfati déclarent:
« En revanche, les supposés quill knobs (points d’attache des plumes) observés chez quelques dinosaures candidats à plumes présentant des tubercules sur l’avant-bras montrent davantage de variations de taille, un espacement irrégulier et, au lieu de former une rangée nette le long de l’arrière (côté caudal) de l’ulna, ils sont orientés selon différents angles le long de l’os. Cette dernière caractéristique signifie que, si ces tubercules étaient réellement des quill knobs, les plumes qui y seraient attachées ressortiraient à des endroits inadaptés pour former des ailes, et peut-être même inadaptés pour toute autre fonction. »
On pourrait aussi évoquer le problème de la queue lourde du théropode, très incongrue pour devenir une queue légère d’oiseau adaptée au vol.

La queue est un vrai empennage d’avion chez l’oiseau. Elle joue un rôle de stabilisation en générant un couple qui équilibre le corps et règle l’angle d’attaque (surtout en vol lent ou avec des charges). Elle permet aussi de commander la trajectoire car en l’écartant/asymétrisant, l’oiseau contrôle tangage, lacet et un peu le roulis, les virages serrés, les corrections fines et le vol dans le vent.
La surface portante supplémentaire abaisse la vitesse de décrochage et la queue repliée permet une traînée minimale pour le vol rapide. Il y a également un rôle de freinage/atterrissage, il y a le grand cabrage final (ou l’arrondi) de la queue pour freiner, stabiliser l’arrondi et poser précisément.
Il y a encore le rôle du décollage & manœuvres près du sol qui aide à arracher le nez3 et à garder le contrôle à très basse vitesse.
Les oiseaux lissent leurs plumes

Les oiseaux lissent leurs plumes pour:
- Ré-emboîter les barbules: en pinçant la plume, le bec « re-zippe » les petits crochets (barbicelles) ce qui perme de retrouver la portance et le silence de vol.
- Étaler l’huile de la glande uropygienne (près de la queue) ce qui permet imperméabilisation, souplesse, protection UV et antimicrobienne/antifongique.
- Éliminer poussières et parasites (poux des plumes, acariens) et retirer les gaines des plumes en croissance.
- Thermorégulation & signal: un plumage propre isole mieux et garde les couleurs/irisations pour la sélection sexuelle.
Le bec est adapté pour faire le travail de lissage. Des pointes fines ou légèrement crochues (passereaux, perroquets) permettent de « peigner » et de pincer les barbules. Une grande dextérité est observée grâce à la langue et à des mécanorécepteurs au bout du bec pour un contrôle très précis.
Sans lissage efficace, beaucoup d’oiseaux ne survivent pas longtemps (surtout en milieu froid ou aquatique). C’est donc, fonctionnellement, indispensable. Quelques jours chez des oiseaux aquatiques peuvent suffire pour entraîner une détresse thermique. Chez les terrestres, on voit rapidement plumage cassant, parasites, baisse de condition, mortalité accrue et échec de reproduction.
Les théropodes maniraptoriens n’avaient ni le bec, ni l’équipement anatomique, ni les preuves comportementales nécessaires pour un lissage des plumes de type avien. Les maniraptoriens typiques (Velociraptor, Deinonychus, Troodon, etc.) avaient des dents en forme de lame, tranchante et robuste, avec une fonction clairement prédatrice contrairement aux dents simples, petites et fines d’Archæoptéryx probablement utilisées pour la préhension plus que la découpe.
Une étude montre qu’Archæoptéryx avait des papilles orales (guidage de matière), une langue osseuse mobile (appareil hyoïdien), et une sensibilité du bout du museau (canaux nerveux).4 Cela permettait probablement de pincer, tirer, repositionner des structures fines. Des animaux actuels dentés (chauves-souris, certains reptiles) utilisent déjà dents + langue + lèvres pour des manipulations fines (toilettage, nettoyage). Un lissage était mécaniquement possible.
Une étude récente indique qu’Archéoptéryx, comme la plupart des oiseaux modernes, utilisait un ensemble de structures coordonnées pour se nourrir.5 Sa langue était ancrée à un os spécialisé lui conférant une grande amplitude de mouvement. Ce mécanisme était associé à des petites protubérances sensibles dans sa bouche, appelées papilles orales, qui guidaient les aliments dans la gorge. Aucun dinosaure ne possède ces caractéristiques.

Le vol optimisé d’Archæoptéryx
Les plumes d’Archaeopteryx, dès les premiers spécimens découverts, sont des plumes pennées asymétriques, identiques à celles des oiseaux modernes, avec un rachis, des barbules et des crochets adaptés au vol.

Des études aérodynamiques (comme Carney et al, 2012) ont montré que les plumes d’Archæoptéryx sont déjà optimisées pour le vol battu et le contrôle de portance, cela est bien plus complexe qu’une étape de « transition ». Cela implique que la machinerie génétique et morphologique des plumes était déjà complète dès son apparition dans le registre fossile.
Si Archaeopteryx est censé être un « prototype », pourquoi ses plumes sont-elles aussi développées que celles d’oiseaux modernes? On ne voit pas de séquence progressive de « proto-plumes » rudimentaires vers des plumes pleinement fonctionnelles (elles apparaissent d’un coup).
Il n’existe pas de fossiles montrant des étapes intermédiaires entre les écailles reptiliennes et les plumes complexes d’Archæoptéryx. L’apparition soudaine de plumes complètes dans les archives fossiles s’aligne avec le paradigme créationniste.

Les évolutionnistes répondent souvent en citant l’expérience de Cooper & Milinkovitch qui a montré que chez l’embryon de poulet, en modulant transitoirement l’activation du gène Sonic hedgehog, une écaille peut se transformer en plume. La conclusion est que si une simple variation dans la régulation suffit à « transformer » une écaille en plume, il n’y a pas de « fossile mi-plume mi-écaille » à attendre, les transitions se font par régulation et pas par structure intermédiaire. On s’éloigne bien sûr du récit évolutif gradualiste initial mais considérons l’idée.
Les poulets, comme tous les oiseaux modernes, ont des plumes, mais aussi des zones recouvertes d’écailles.
Sur les pattes et les doigts, on trouve des écailles cornées (kératine), très semblables à celles des reptiles. Ce sont des écailles « aviennes » qui ne sont pas identiques à celles des reptiles: elles se forment à partir des mêmes placodes embryonnaires que les plumes, mais le programme génétique module le développement pour donner une écaille au lieu d’une plume. On parle de scutelles (grandes écailles recouvrant la face antérieure des tarses) et de scutellules (plus petites, couvrant les doigts). C’est sur ces écailles de pattes que l’expérience de Milinkovitch (2023) a injecté le signal Sonic Hedgehog (Shh):
- Normalement ces placodes donnent des écailles sur les pattes.
- Avec l’activation forcée de Shh, elles ont produit des plumes normales, complètes.
Cela démontre que le code pour produire des plumes est déjà présent partout, mais seulement activé dans certaines zones. Le programme était déjà là. Les poulets possèdent déjà toutes les instructions génétiques pour produire des plumes. L’expérience ne crée rien de nouveau, elle ne fait que détourner un programme existant. Si les dinosaures n’avaient pas ce programme, une telle manipulation n’aurait jamais produit de plumes.
C’est une fausse démonstration de l’évolution. Présenter cette expérience comme « preuve » de la transition écaille/plume est trompeur car on utilise un animal comme la poule qui a déjà le code pour les plumes. Faire apparaître des plumes sur une poule n’a rien de surprenant.
C’est la même chose qu’avec les expériences anciennes où on utilisait de la vitamine A pour induire des « plumes » sur des embryons de poulet. Cela ne prouvait rien sur l’origine des plumes, seulement que le programme génétique était flexible.
On observe plutôt une sophistication qui pointe vers un Concepteur. La régulation Shh est tellement fine qu’une petite perturbation tue l’embryon. Cela montre que nous sommes devant un système hautement intégré et réglé, ce qui est mieux expliqué par la conception intelligente que par des mutations aléatoires.
L’expérience ne démontre pas que les plumes ont évolué à partir d’écailles, mais que les poulets possèdent déjà un logiciel génétique sophistiqué, modulable mais fragile. Cela correspond davantage à un programme intentionnel qu’à un bricolage aveugle.
Feo, Field & Prum (2015) est une étude morphologique évolutionniste essayant de démontrer que la géométrie des barbes de plumes asymétriques reflèterait une étape intermédiaire dans l’évolution du vol avien.
Les plumes de vol asymétriques sont une des caractéristiques les plus distinctives des oiseaux actuels. Archæoptéryx présente le même de type plumes asymétriques qui sont quasi indiscernables de celles des oiseaux modernes.
L’étude de Feo et de ses collègues ne contredisent pas cela, ils parlent d’un vol moins sophistiqué que celui des oiseaux modernes, leur image est de comparer l’avion des frères Wright avec un jet fighter moderne. Toutefois si Archéoptéryx est un oiseau d’un autre clade avec ses différences et un autre type de vol c’est normal.
Les différences de géométrie (angle des barbes, rapport longueur/épaisseur, densité, etc.) reflètent une variation fonctionnelle parmi les oiseaux et non obligatoirement une étape évolutive. On observe aujourd’hui des variations comparables entre oiseaux modernes selon le mode de vol avec les faucons qui ont des plumes très asymétriques pour le vol rapide et plané, les hiboux qui ont des barbes plus souples, des bords frangés pour le vol silencieux et les faisans/dindes qui ont des plumes épaisses et arrondies pour le vol court et le décollage rapide. Ces différences ne font pas d’eux des transitions mais des adaptations à des usages spécifiques.
Les oiseaux volent, mais de manières très différentes, selon des niches et des designs adaptés.
Bhullar et al. (2015) est une étude de biologie évolutive du développement qui propose un mécanisme moléculaire expliquant l’origine évolutive du bec et du palais des oiseaux à partir de leurs ancêtres reptiliens. Les manipulations génétiques en laboratoire produisent des déformations et pas de véritable « évolution » d’un bec. Cela montre que l’information était déjà présente dans le génome et non pas qu’un mécanisme aveugle a créé une innovation majeure.
Rashid et al. (2014) est une étude sur la transformation évolutive de la queue des dinosaures vers les oiseaux, en lien avec les changements de développement embryonnaire. Les différences de queue (longue chez les dinosaures, pygostyle chez les oiseaux) sont des catégories distinctes. Les fossiles montrent deux types bien définis, non pas une série continue d’intermédiaires.
Frank Sherwin résume le vol d’Archaeopteryx comme suit:6
« Le paléontologue évolutionniste Alan Feduccia a souligné que les plumes d’Archaeopteryx sont complètement aérodynamiques dans leur structure, ce qui indique le vol. De plus, Archaeopteryx possédait une “fourchette” (clavicule en V) suffisamment robuste pour l’attache musculaire et la capacité de vol. En outre, l’os du pubis chez Archaeopteryx ressemble à celui des oiseaux actuels et servait probablement de site d’attache pour les muscles suprapubiens, qui sont conçus pour aider à la ventilation pulmonaire en bougeant la queue pendant le repos. En revanche, l’os du pubis des crocodiles et des dinosaures bipèdes fonctionne comme un site d’attache pour les muscles du diaphragme.
La question du vol a été tranchée après que des scientifiques aient étudié la géométrie en coupe transversale des os des ailes de trois spécimens d’Archaeopteryx. Ils ont conclu qu’Archaeopteryx volait comme un faisan, avec des ascensions rapides, un temps de vol court, et la course comme moyen d’échapper au danger.
« Nos analyses révèlent que l’architecture des os des ailes d’Archaeopteryx présente systématiquement une combinaison de propriétés géométriques en coupe transversale partagées de manière unique avec les oiseaux volants, en particulier ceux qui utilisent occasionnellement le battement d’ailes sur de courtes distances. En effet, Archaeopteryx semble avoir volé en battant des ailes par petites rafales. »
Le paléontologue évolutionniste Steve Brusatte a déclaré : *« Je pense que l’affaire est désormais close… Archaeopteryx était capable d’au moins de courtes rafales de vol battu. »*«
Le système respiratoire
Les fossiles d’Archaeopteryx montrent des cavités pneumatiques dans les os, très similaires à celles des oiseaux modernes, une structure thoracique compatible avec le fonctionnement des sacs aériens.
Certaines études (Ruben et al., 1997 ; Feduccia, 1999) suggèrent que les théropodes n’avaient pas un système respiratoire adapté au vol contrairement à Archæoptéryx. Cela indique qu’ils avaient déjà un système respiratoire aviaire complet et non intermédiaire.
Les reptiles (comme les dinosaures théropodes) avaient des poumons sacculaires (comme nous) avec l’air qui entre et ressort par le même chemin (ventilation bidirectionnelle). Cela a des limites car il n’y a pas d’échange optimal et l’air frais se mélange à l’air vicié.
Les oiseaux modernes ont système à flux d’air unidirectionnel, grâce à des sacs aériens reliés aux poumons. Les avantages sont un apport constant d’air riche en O₂, même à l’expiration. Cela permet un métabolisme très élevé qui est indispensable pour le vol actif. C’est un système radicalement différent, qui ne peut pas être fonctionnel à moitié.
Si Archæoptéryx descend des dinosaures théropodes, il aurait fallu une transformation complète du système respiratoire pour passer d’un système bidirectionnel reptilien à un système unidirectionnel aviaire mais cela exige des changements radicaux (structure des poumons, os, sacs aériens, sternum, circulation sanguine). Or, aucun fossile transitionnel ne montre ce passage périlleux.
Les évolutionnistes rétorquent parfois avec le système respiratoire des alligators, lesquels ont également un circuit unidirectionnel mais leurs poumons sont expansibles avec un muscle diaphragmatique (non rigides comme les oiseaux), ils n’ont pas de sacs aériens et ont une ventilation par piston hépatique (comme les crocodiles). Cela est bien différent du système respiratoire des oiseaux avec poumons rigides + sacs aériens + ventilation par sternum.
C’est un exemple d’irréductible complexité physiologique, sans système complet, il n’y a pas de respiration et donc pas de survie.

Le système respiratoire d’Archæoptéryx est déjà celui des oiseaux modernes, fondé sur les sacs aériens et la ventilation unidirectionnelle. Les reptiles ont quant à eux un système totalement différent. Ce saut qualitatif radical est un argument fort pour la création car il ne peut pas résulter de petites étapes graduelles, cela renforce l’idée d’un plan de conception dès l’origine.
Ce point de vue est confirmé par des non-créationnistes, voici une citation d’Ann Gibbons (journaliste scientifique) au sujet de l’évolution des dinosaures au oiseaux:
«Voici la première preuve dans les tissus mous que les théropodes avaient le même type de compartimentation des poumons, du foie et des intestins que l’on trouverait chez un crocodile – et non chez un oiseau.
John Ruben (paléontologue de vertébrés) utilise cette preuve pulmonaire pour argumenter non seulement que les dinosaures étaient incapables des taux élevés d’échange de gaz nécessaires pour l’homéothermie (particulière aux animaux à sang chaud comme les oiseaux), mais aussi que leurs poumons semblables à des soufflets n’auraient pas pu évoluer vers les poumons « haute performance » des oiseaux modernes»
“Ruben, J.A., cité dans Ann Gibbons, Lung fossils suggest dinos breathed in cold blood, Science 278(5341):1229–1230, 1997.”
John Ruben déclare encore:
« Récemment, la sagesse conventionnelle a soutenu que les oiseaux descendent directement des dinosaures théropodes. Cependant, le maintien apparemment inébranlable de la ventilation pulmonaire par piston hépatique (diaphragmatique) chez les théropodes tout au long du Mésozoïque pose des problèmes fondamentaux pour une telle relation.
Les premiers stades de la dérivation du système de sacs aériens abdominaux aviaires à partir d’un ancêtre ventilant par diaphragme auraient nécessité une sélection en faveur d’une hernie diaphragmatique chez les taxons transitionnels entre théropodes et oiseaux. Une telle condition invalidante aurait immédiatement compromis l’ensemble de l’appareil ventilatoire pulmonaire et il semble improbable qu’elle ait représenté un quelconque avantage sélectif. »
Ruben, J.A. et al. 1997. Lung Structure and Ventilation in Theropod Dinosaurs and Early Birds. Science. 278 (5341): 1269.
Une partie de ce système respiratoire unique (des poumons fixes + un réseau de sacs aériens qui assurent une ventilation unidirectionnelle), le subpectoral diverticulum (SPD), est une extension placée entre les muscles pectoraux principaux (ceux qui actionnent les ailes).
Quand il se gonfle, il optimise l’effet de levier des muscles, stabilise les ailes en vol plané ou en vol battu, et réduit la dépense énergétique. L’étude de Schachner et al. (2024) a montré que cette structure est présente chez plusieurs groupes d’oiseaux très différents.
John Ruben déclare encore dans une étude de 2009:
« L’absence probable d’une fonction pulmonaire de type aviaire chez les théropodes est incompatible avec l’hypothèse d’une anatomie cardiovasculaire plus sophistiquée que celle des crocodiliens modernes. »
Cardio-pulmonary anatomy in theropod dinosaurs: Implications from extant archosaurs

Les chercheurs concluent que le SPD aurait évolué indépendamment au moins 7 fois dans des lignées d’oiseaux séparées (convergence évolutive). Autrement dit, le même système complexe, impliquant des interactions précises entre sacs aériens, muscles et mécanique de vol, serait apparu par hasard à plusieurs reprises.
C’est exactement le même problème que pour les yeux, les ailes ou les organes sensoriels complexes: on invoque la convergence parce que l’arbre évolutif ne permet pas de postuler un ancêtre commun. Mais chaque « apparition indépendante » signifie en fait que l’évolution doit réinventer le même mécanisme complexe plusieurs fois, ce qui devient de plus en plus improbable.
Dans un modèle de concepteur commun, ce constat est attendu:
- Un même ingénieur peut réutiliser un système optimisé (ici le SPD) dans différents « modèles » d’oiseaux.
- L’économie d’énergie est cruciale pour le vol, ce type de design transversal est parfaitement rationnel dans une perspective d’ingénierie.
Au lieu de parler de « convergences évolutives » improbables, il est plus cohérent de parler de réutilisation de modules fonctionnels. Cette découverte illustre une tendance claire:
- Pour l’évolution, chaque nouvel exemple de convergence complique l’histoire et oblige à multiplier les « apparitions indépendantes ».
- Pour la création, au contraire, chaque convergence renforce l’idée d’un plan de conception optimisé et modulaire, qui explique pourquoi on retrouve des solutions identiques dans des groupes très éloignés.
Smith et al. (2015), The Auk est une réponse critique à l’article de Feduccia contestant l’origine dinosaurienne des oiseaux, défendant au contraire le consensus paléontologique d’un lien étroit entre oiseaux et dinosaures théropodes.
Le fait que des évolutionnistes débattent encore de l’origine des oiseaux (dinosaures vs origine indépendante) montre que le dossier fossile n’est pas clair. Ce désaccord démontre que le lien oiseau–dinosaure est basé sur des interprétations et non des preuves directes.
Les dinosaures: des animaux à « sang froid »?
Depuis longtemps, les scientifiques se demandent comment des reptiles à sang froid comme les dinosaures ont pu être aussi actifs, rapides et dominants. Bien que les fossiles montrent de fortes ressemblances avec les reptiles actuels, une hypothèse évolutionniste populaire affirme que les dinosaures auraient été « à sang-chaud », proches des oiseaux, et donc très différents des reptiles modernes (une idée contre-intuitive). De nouvelles recherches en physiologie et génétique des reptiles vivants apportent cependant un éclairage différent.7
De nombreux scientifiques évolutionnistes cherchent à montrer que les dinosaures étaient endothermes (à sang chaud) afin de les rapprocher biologiquement des oiseaux, et de soutenir l’idée que « les oiseaux sont des dinosaures » et que les dinosaures ne se seraient donc jamais vraiment éteints. Les preuves avancées sont toutefois exagérées ou mal interprétées.8
Déterminer si un animal éteint était endotherme (production interne de chaleur) ou ectotherme (dépendant de la chaleur externe), est extrêmement difficile, car cela nécessite d’étudier les organes, les systèmes respiratoires etc… qui ne sont pas disponibles chez les fossiles.
Les partisans des dinosaures à sang chaud soulignent que les os de dinosaures sont très vascularisés, comme chez les mammifères mais la vascularisation indique surtout un niveau d’activité élevé et pas nécessairement un métabolisme endotherme.
Une étude de Tomasz Owerkowicz9 montre que des animaux actifs peuvent avoir des os très vascularisés sans être à sang chaud.
Les voies nasales représentent un test plus pertinent. Les cornets nasaux sont un bien meilleur indicateur car 99 % des animaux à sang chaud en possèdent, ils réduisent les pertes de chaleur et d’eau lors de la respiration. Les scans CT réalisés par John Ruben10 sur plusieurs crânes de dinosaures montrent des cavités nasales étroites, similaires à celles des crocodiles et autres reptiles à sang froid.
D’autres indices vont dans le même sens. Scipionyx, un petit dinosaure souvent présenté comme proche des oiseaux possédait un système respiratoire similaire aux crocodiles et pas de poumon à sacs aériens avec flux unidirectionnel comme chez les oiseaux. L’attache des intestins confirme l’absence d’un système respiratoire avien. Ces données vont à l’encontre du scénario théropode-oiseau.
Certains chercheurs proposent que les dinosaures étaient mésothermes (comme le thon ou la tortue luth), et donc capables d’élever leur température par l’activité, mais incapables de la maintenir indépendamment de l’environnement. Cela rapproche encore les dinosaures des ectothermes et pas des oiseaux.
Rien n’oblige scientifiquement à considérer les dinosaures comme à sang chaud, les données disponibles soutiennent plutôt l’idée de dinosaures reptiliens à sang froid, dont la physiologie aurait contribué à leur extinction après le Déluge (ère glaciaire).
Les évolutionnistes ont besoin de dinosaures à sang chaud pour maintenir le lien dinosaures-oiseaux, il est cependant difficile de faire dériver un oiseau endotherme d’un reptile ectotherme, ce besoin théorique pousse à défendre une position non démontrée.
Les preuves souvent citées pour faire des dinosaures des animaux à sang chaud ne sont ni décisives ni cohérentes, et plusieurs données anatomiques les rapprochent davantage des reptiles que des oiseaux, ce qui affaiblit le scénario évolutif dinosaures-oiseaux.
Les fossiles de dinosaures présentent plusieurs caractéristiques typiquement reptiliennes: des cavités nasales étroites, sans cornets, contrairement aux mammifères à sang chaud, et similaires à celles des reptiles actuels.
Mais aussi un système respiratoire de type reptilien, avec une ventilation pulmonaire faible au repos. Les fossiles de tissus mous abdominaux chez des théropodes montrent un système respiratoire comparable à celui des crocodiles. Ces éléments vont à l’encontre d’un métabolisme avien.
Les chercheurs ont proposé que de nombreux dinosaures avaient un métabolisme bas au repos, mais pouvaient augmenter rapidement leur ventilation pulmonaire pour courir, attaquer et se défendre. Ce modèle expliquerait leur grande activité ponctuelle, sans nécessiter un métabolisme à sang chaud permanent. Il résolverait aussi un problème énergétique majeur: un dinosaure géant à sang chaud n’aurait pas pu manger assez pour survivre, alors qu’un ectotherme flexible, oui.
Des études récentes confirment que ce type de métabolisme existe aujourd’hui. Le dragon de Komodo, le plus grand lézard terrestre vivant (jusqu’à 10 m / 300 kg), est un prédateur très actif. Les recherches montrent qu’il peut augmenter son métabolisme à des niveaux quasi mammaliens, sur de courtes ou moyennes durées (sprints, poursuites), tout en restant fondamentalement ectotherme. Le séquençage de son génome a révélé des adaptations génétiques spécifiques liées à cette performance.
Une étude comparative entre T. rex et des alligators vivants a montré que tous deux possèdent deux grandes ouvertures au sommet du crâne. Chez les alligators ces cavités contiennent des vaisseaux sanguins spécialisés, impliqués dans la thermorégulation. L’imagerie thermique révèle une activité métabolique variable selon la température externe, faisant partie d’un système circulatoire de régulation thermique.
Ces structures, longtemps interprétées comme des muscles chez les dinosaures, seraient en réalité thermorégulatrices, comme chez les reptiles actuels.
Les similitudes croissantes entre dinosaures fossiles et reptiles vivants (crocodiles, varans), confirment que les dinosaures étaient de véritables reptiles à sang froid, dotés d’une physiologie hautement optimisée, capables de performances élevées sans être endothermes.
Ces découvertes affaiblissent le lien dinosaures-oiseaux et mettent en valeur une conception adaptative remarquable dans le monde vivant. Les dinosaures n’avaient pas besoin d’être à sang chaud pour être actifs car comme certains reptiles modernes, ils combinaient un métabolisme bas au repos avec des pics de performance élevés, ce qui les identifie comme de véritables reptiles ectothermes très bien conçus.
Chez l’oiseau l’endothermie (production interne de chaleur) est vitale car le vol est énergétiquement exorbitant. Le vol battu est le mode de locomotion le plus coûteux du règne animal. Il nécessite des battements rapides et continus, des muscles pectoraux hypertrophiés, un métabolisme 2 à 3 fois plus élevé que chez un mammifère de même masse.
Sans production interne constante de chaleur, les muscles perdent en puissance, se refroidissent rapidement et deviennent inefficaces. Un animal ectotherme (à sang froid) ne peut pas soutenir un vol battu prolongé.
Les réactions biochimiques ont une plage thermique étroite. Chez les oiseaux on observe température corporelle d’environ 40–42 °C (plus élevée que chez les mammifères), et des enzymes optimisées pour ce régime thermique élevé. Sans endothermie la contraction musculaire devient lente, la coordination fine du vol s’effondre, le décollage devient impossible. Le vol exige une thermostabilité et non juste des « pics d’activité ».
Le vol implique une énorme perte de chaleur. Le paradoxe du vol est que malgré la production massive de chaleur par les muscles, il y a une perte de chaleur extrême due à l’air, à l’altitude, à la convection. Les oiseaux compensent par une endothermie élevée, des plumes isolantes et une régulation vasculaire fine. Un ectotherme volant se refroidirait plus vite qu’il ne pourrait produire de l’énergie.
L’endothermie permet aux oiseaux de voler à l’aube, la nuit, en hiver, de migrer sur des milliers de kilomètres, d’occuper des niches arctiques, montagnardes, océaniques. Sans endothermie il n’y a pas de migration longue distance, pas de vol nocturne, pas de vie active en climat froid.
Les oiseaux ont des poumons rigides, des sacs aériens, un flux d’air unidirectionnel. Ce système maximise l’oxygénation, alimente un métabolisme extrêmement élevé et dissipe la chaleur excédentaire. Ce système n’a de sens que couplé à l’endothermie.
Les oiseaux incubent leurs œufs à température constante, indépendamment de l’environnement. Sans endothermie il y aurait une incubation instable, un développement embryonnaire compromis et une dépendance extrême au climat.
Une étude majeure publiée dans PNAS (2017) par Gregory Erickson et al., a d’ailleurs mesuré directement la durée d’incubation des œufs de dinosaures.11 C’est un point crucial, parce que le temps d’incubation est fortement lié au métabolisme et c’est un critère clé pour savoir si un animal est reptilien (ectotherme) ou avien/mammalien (endotherme).
Les chercheurs ont étudié des dents embryonnaires fossiles chez:
- Protoceratops andrewsi,
- Hypacrosaurus stebingeri.
Ils ont découvert dans ces dents des lignes de croissance quotidiennes (von Ebner lines), équivalentes aux cernes d’un arbre, déjà connues chez les reptiles et mammifères actuels. Chaque ligne est égale à 1 jour de développement embryonnaire. En les comptant, on obtient l’âge exact de l’embryon à l’éclosion. C’est une mesure directe et non un modèle théorique.
Les résultats ont surpris les auteurs eux-mêmes:
| Espèce | Incubation attendue (oiseau) | Incubation mesurée |
|---|---|---|
| Protoceratops | 40 jours | 83 jours |
| Hypacrosaurus | 82 jours | 171 jours |
C’est environ 2 fois plus long que chez les oiseaux de taille comparable.
- Chez les animaux ovipares, les oiseaux endothermes ont une incubation courte, un métabolisme embryonnaire élevé et des œufs peu nombreux mais gros.
- Chez les reptiles ectothermes on observe une incubation longue, un métabolisme lent et des pontes nombreuses.
Or les dinosaures montrent des grandes pontes (20–30 œufs), une incubation lente, un métabolisme embryonnaire bas. Tous les marqueurs vont du côté reptilien, et non du côté avien.
L’incubation lente implique une production d’énergie faible par unité de temps, incompatible avec l’endothermie, un développement rapide type oiseau. Même les auteurs de l’étude reconnaissent implicitement que les dinosaures se situent hors du modèle avien. Mais comme le souligne Clarey12, ils évitent d’en tirer une conclusion métabolique claire car cela entrerait en conflit avec le récit théropode-oiseau.
Les oiseaux se distinguent par une incubation rapide, un contrôle thermique parental, des œufs peu nombreux. Les dinosaures étudiés montrent l’inverse. Si les dinosaures étaient réellement des oiseaux: leurs œufs auraient dû se comporter comme des œufs d’oiseaux, ce qui n’est pas le cas.
Cette étude démontre solidement que les œufs de dinosaures se comportaient comme ceux des reptiles et pas comme ceux des oiseaux. Cela démontre le métabolisme embryonnaire lent des dinosaures, leur incubation longue et leur stratégie reproductive non avienne. Ce sont des données concrètes, difficiles à concilier avec l’idée évolutive que les « oiseaux sont des dinosaures ».
Les cuisses internes des oiseaux
Une étude de l’Oregon State University, dans la lignée de celles de John Ruben, a aggravé le problème de la transformation d’un système respiratoire reptilien à celui d’un oiseau comme Archæoptéryx. Il s’avère que la configuration des os d’oiseaux est essentielle pour leur respiration.
Chez les dinosaures théropodes (comme Velociraptor et Deinonychus):
- Les cuisses sont externes au tronc.
- Le fémur est placé en dehors de la cage thoracique.
- La locomotion se fait avec un mouvement de la hanche vers l’extérieur, comme chez les reptiles modernes.
- Cela implique une respiration de type reptilien, souvent par piston hépatique, comparable aux crocodiles.
Chez les oiseaux modernes:
- Les cuisses sont internes, enchâssées dans le tronc.
- Le fémur reste en position quasi horizontale et fixe à l’intérieur du corps.
- La locomotion vient surtout du genou et du tarse et pas de la hanche.
- Cette configuration permet d’intégrer le système respiratoire avien unique constitué de poumons fixes avec sacs aériens, ce qui favorise un flux d’air unidirectionnel.
- Si les cuisses sortaient du tronc comme chez les dinos, ce système serait compromis.

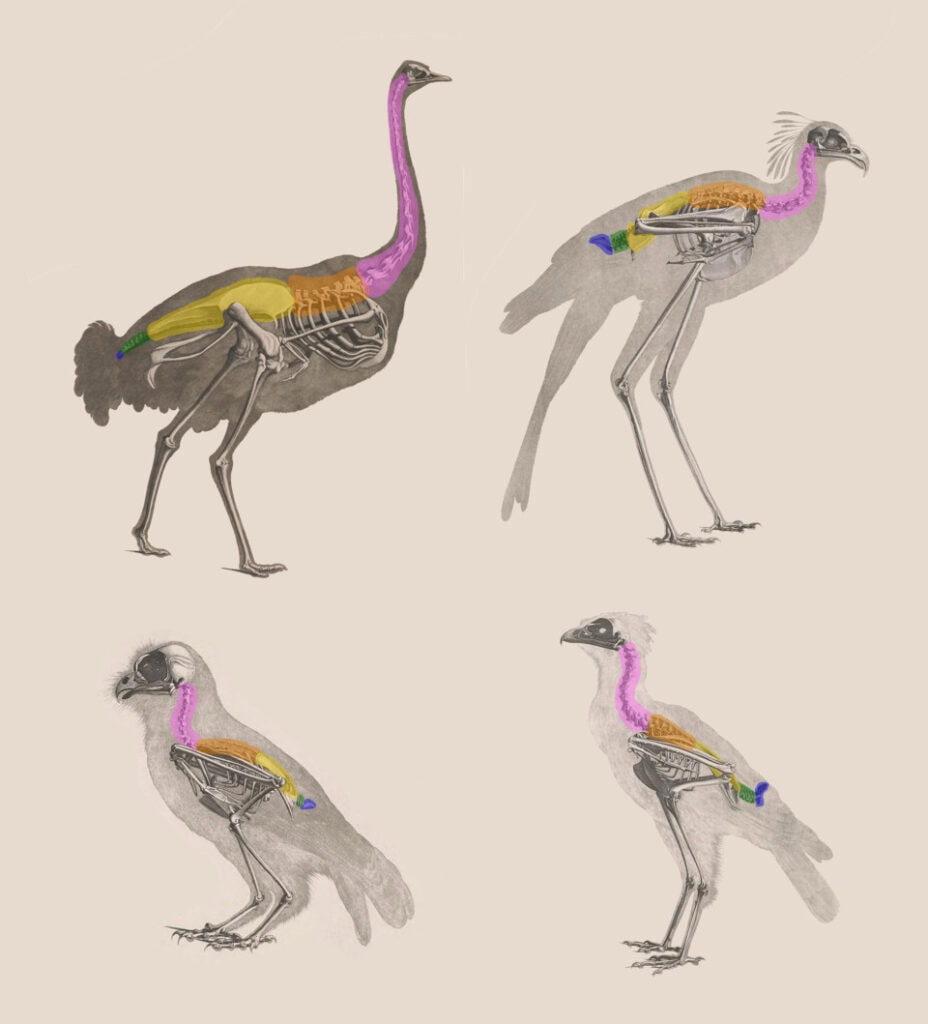
Passer de cuisses externes de type reptile à cuisses internes de type oiseau ne peut pas se faire par « petites étapes » adaptatives. Cela impliquerait une reconstruction complète de la hanche, du bassin, des muscles locomoteurs et du système respiratoire. Une transition intermédiaire serait inviable par la perte de mobilité ou l’incapacité respiratoire.
Ruben (1997, Science) a insisté que cette différence est « fondamentale et non triviale », rendant peu plausible un scénario de continuité entre dinosaures théropodes et oiseaux.
Cette distinction appuie l’idée que les oiseaux et les dinosaures sont deux groupes distincts, créés séparément. Les oiseaux ont été conçus dès le départ avec une intégration unique comprenant une respiration ultraperformante + une anatomie locomotrice adaptée au vol. Les dinosaures ont quant à eux une respiration reptilienne, qui ne peut pas « évoluer » graduellement vers le système avien sans étapes létales.
L’un des coauteurs de l’étude, la zoologiste de l’OSU Devon Quick, a déclaré dans un communiqué de presse de l’université:
« C’est vraiment étrange que personne ne s’en soit aperçu plus tôt. La position du fémur et des muscles chez les oiseaux est cruciale pour le fonctionnement de leurs poumons, ce qui leur donne en retour la capacité pulmonaire suffisante pour voler. »
Discovery raises new doubts about dinosaur-bird links. Oregon State University press, June 9, 2009.
Il a également déclaré:
« Chez les oiseaux, le fonctionnement normal des poumons requiert de vastes sacs aériens ventilés par un sternum expansif et des côtes spécialement articulées… [ainsi qu’un] complexe fémoral-cuisse spécialisé. »
Quick, D.E. and Ruben, J.A. Cardio-pulmonary anatomy in theropod dinosaurs: Implications from extant archosaurs. Journal of Morphology. 20 mai, 2009.
Puis:
« L’absence probable d’une fonction pulmonaire semblable à celle des oiseaux chez les théropodes est incompatible avec les suggestions d’une anatomie cardiovasculaire plus sophistiquée que celle des crocodiliens modernes. »
Quick, D.E. and Ruben, J.A. Cardio-pulmonary anatomy in theropod dinosaurs: Implications from extant archosaurs. Journal of Morphology. 20 mai, 2009.
Brian Thomas de l’Institute of Creation Research déclare:
« Ces caractéristiques [d’oiseau] ne se retrouvent pas dans les fossiles de théropodes, dont les oiseaux seraient censés avoir évolué. Les dinosaures bipèdes avaient une structure et une fonction des pattes significativement différentes. Contrairement aux oiseaux, qui courent à partir du genou, la mobilité des fémurs des théropodes était essentielle à leur mode de locomotion. »
Fixed Bird Thigh Nixes Dino-to-bird Development – 22 juin 2009
L’acétabulum perforé
Chez les dinosaures (théropodes, sauropodes, ornithischiens) le cotyle (acetabulum) est perforé. On parle d’acetabulum ouvert (open hip socket). La tête du fémur s’articule dans un anneau osseux, sans fond osseux complet. La conséquence fonctionnelle est qu’ils ont des membres strictement verticaux sous le corps avec une démarche érigée, efficace pour la locomotion terrestre. C’est un trait clé et unique des dinosaures.
Ce caractère permet de distinguer les dinosaures des reptiles « classiques » à posture étalée (lézards, crocodiliens actuels qui n’ont pas d’acétabulum complètement ouvert).
Chez les oiseaux (Anchiornis, Microraptor et Archæoptéryx inclus) le cotyle est fonctionnellement fermé ou semi-fermé. Le fond est renforcé par la fusion du bassin, le synsacrum (fusion vertèbres + bassin). La tête du fémur est solidement enchâssée. Cela permet une stabilisation extrême du bassin pour le vol, l’atterrissage, la bipédie dynamique et la transmission efficace des forces entre ailes, pattes et tronc.
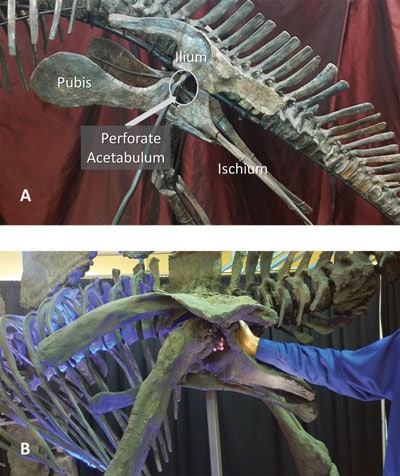
Cotyle perforé chez les dinosaures.
A) Une armature en acier traverse le cotyle perforé d’un fossile original
d’Edmontosaurus juvénile . Les os de la ceinture pelvienne des ornithischiens sont également indiqués.
B) Les doigts de l’auteur (BT) dépassent du cotyle ouvert d’une réplique de fossile
de Stegosaurus adulte (également de l’ordre des Ornithischiens). On notera également les apophyses épineuses caudales proéminentes, points d’insertion des muscles de la queue. Ces apophyses sont absentes chez les oiseaux.
Passer d’un acetabulum ouvert à un bassin fermé et fusionné correspond à un changement d’ingénierie locomotrice, ce n’est pas une simple continuité anatomique. L’acetabulum fermé / renforcé des oiseaux fait partie d’un complexe verrouillé avec bassin fusionné, synsacrum, orientation du fémur, centre de gravité déplacé, lesquels produisent un ensemble indispensable au vol et à l’atterrissage.
Le site du Musée de paléontologie de l’Université de Californie affirme:
« Les dinosaures sont uniques parmi tous les tétrapodes [créatures à quatre membres] par la présence de cet acétabulum perforé. »
Dinosauria: Morphology, University of California Museum of Paleontology, ucmp.berkeley. edu
Les oiseaux (Archæoptéryx et Anchiornis inclus) n’ont pas d’acetabulum pleinement ouvert donc on ne peut pas les appeler « dinosaures ».
Comparatif entre théropodes et oiseaux
| Caractéristique | Théropodes (dinosaures bipèdes) | Oiseaux (modernes & Archaeopteryx) |
|---|---|---|
| Position du fémur (cuisse) | Mobile, détaché du corps | Fixé contre le corps, quasi immobile |
| Mode de locomotion | Propulsion « de la hanche » (comme les humains) | Propulsion « du genou » (cuisse immobile, mouvement du genou et du tarse) |
| Respiration | Pulmonaire type reptilien (souvent supposé « piston hépatique ») | Système de sacs aériens + sternum expansif + côtes articulées |
| Lien entre hanche et respiration | La mobilité du fémur empêche l’intégration des sacs aériens aviaires | La fixation du fémur stabilise le thorax et permet la respiration à flux continu |
| Capacité au vol | Inexistante (aucun système respiratoire assez efficace) | Optimisée pour le vol grâce au système unidirectionnel |
| Analogie moderne | Comme les humains ou crocodiles (hanche mobile) | Unique dans le règne animal, nécessaire au vol soutenu |
Sinosauropteryx
Sinosauropteryx est un petit dinosaure théropode, daté du Crétacé inférieur (125 Ma). Il existe à son sujet des fossiles préservés avec des structures fibreuses autour du corps qui sont interprétées comme des proto-plumes. Ces proto-plumes seraient les ancêtres des plumes vraies d’Archaeopteryx (150 Ma). Il serait une preuve que les plumes sont apparues d’abord comme structures isolantes, puis adaptées au vol.
Les structures observées sont filamenteuses, elles ne disposent aucunement des caractéristiques des plumes vraies (rachis, barbes, barbules, crochets). Plusieurs paléontologues séculiers (même non-créationnistes, ex. Feduccia, 1999) ont soutenu qu’il s’agissait plutôt de fibres de collagène dégradées, provenant de la peau ou du tissu sous-cutané en décomposition. Rien ne prouve donc qu’il s’agissait de plumes ou de précurseurs de plumes.
Bibliquement, des dinosaures à plumes ne poseraient pas de problèmes, dans le cas de Sinosauropteryx cependant, l’identification de plumes n’est pas convaincante.
Archæoptéryx est daté 25 millions d’années avant Sinosauropteryx (Jurassique vs Crétacé) selon le référentiel évolutif. Or, Archæoptéryx possède déjà des plumes modernes et optimisées pour le vol. Comment un dinosaure postérieur, avec de simples filaments non fonctionnels, pourrait-il être l’ancêtre d’un oiseau déjà parfaitement équipé?
Même d’un point de vue évolutif, cela n’a pas de sens mais les évolutionnistes indiquent que le grand-père a pu survivre plus longtemps que son petit-fils…
Les oiseaux nécessitent un ensemble complet (plumes, squelette allégé, muscles, système respiratoire aviaire). Sinosauropteryx n’a aucune de ces caractéristiques car il n’avait ni ailes ni capacités de vol. Le relier à Archæoptéryx repose uniquement sur une interprétation spéculative des filaments.
Du point de vue créationniste:
- Archaeopteryx était un oiseau complet, avec plumes et capacités de vol.
- Sinosauropteryx était un dinosaure terrestre, sans plumes.
- Les deux font partie de baramins distincts (groupes créés séparés).
« Il manquait le poumon avien unique. À la place, il possédait un système respiratoire de type crocodilien, dit ‘hepatic piston’, fonctionnant selon la méthode des soufflets reptiliens normaux. John Ruben, expert en physiologie respiratoire à l’Oregon State University à Corvallis, a conclu que les “poumons en forme de soufflet de Sinosauropteryx ne peuvent pas avoir évolué en poumons haute performance des oiseaux modernes.” »
Ruben, J.A., quoted in Ann Gibbons, Lung fossils suggest dinos breathed in cold blood, Science 278(5341):1229–1230, 1997.
Selon John Ruben, le système respiratoire de type reptilien chez Sinosauropteryx n’a aucun chemin plausible pour se transformer graduellement en poumons aviaires avec sacs aériens et ventilation unidirectionnelle.
Le Professeur Theagarten Lingham-Soliar, de l’Université du KwaZulu Natal en Afrique du Sud, a indiqué que les filaments faisaient partie:
« d’une structure clairement unifiée, c’est-à-dire une collerette ou une crête dressée recouvrant le cou, le dos et la queue de Sinosauropteryx … plutôt que de proto-plumes individuelles ».
Lingham-Soliar T., The evolution of the feather: Sinosauropteryx, life, death, and preservation of an alleged feathered dinosaur, J. Ornithol 153(3):699–711, 2012.
Mike Oard écrit:
« Bien qu’il soit possible que les structures tégumentaires observées soient des plumes véritables dégradées, cette déduction pose plusieurs problèmes. Feduccia et d’autres ont analysé en détail les affirmations concernant les dinosaures à plumes de Chine. Ils ont mis en évidence des données expérimentales et fossiles suggérant que les fibres observées le long de l’extérieur du squelette de Sinosauropteryx et d’autres spécimens s’expliquent mieux comme des fibres de collagène faisant simplement partie de la peau du dinosaure.
Nous savons également qu’il existe plusieurs dinosaures sans lien évolutif supposé avec les oiseaux qui présentent eux aussi ces fibres. L’un d’eux est Psittacosaurus, un dinosaure à cornes semblable aux cératopsiens. Un autre est un nouveau dinosaure “à hanches d’oiseau” découvert en Chine qui, soit dit en passant, a étendu l’intervalle stratigraphique de ce type particulier de dinosaure jusqu’à une période plus récente d’au moins 60 millions d’années, illustrant, au minimum, que de nouvelles découvertes peuvent imposer des révisions théoriques. Les oiseaux ne sont pas censés avoir évolué à partir de dinosaures à hanches d’oiseau [Ornithischia], mais à partir de dinosaures à hanches de lézard [Saurischia], ce qui explique pourquoi la découverte de structures tégumentaires chez un dinosaure à hanches d’oiseau est si significative.
D’autres types d’animaux fossiles présentent également ces fibres de collagène en forme de soies, comme un ptérosaure, un ichthyosaure, ainsi que d’autres reptiles et dinosaures. Même des fibres de collagène provenant d’un dauphin mort étaient similaires aux proto-plumes revendiquées en Chine. Ainsi, ces structures fibreuses ne peuvent pas être considérées comme propres aux dinosaures situés dans la chaîne évolutive menant aux oiseaux, ce qui soutient également la conclusion de Feduccia et al. selon laquelle il s’agit simplement de fibres de collagène. »
Confuciusornis
Confuciusornis est un oiseau avec des plumes parfaitement formées au même moment que des dinosaures plumeux supposés (Sinosauropteryx), cela montre qu’il n’y a pas de transition mais des catégories bien séparées.
Comme Archaeopteryx, Confuciusornis montre un oiseau déjà pleinement formé. Il n’y a pas de trace de « proto-plumes », ses plumes sont modernes, fonctionnelles, optimisées. Il est daté par les évolutionnistes du Crétacé inférieur (125 Ma), donc supposément 25 millions d’années plus jeune qu’Archæoptéryx (150 Ma). Il a la taille d’un pigeon et est considéré comme le plus ancien oiseau doté d’un bec sans dents.
En plus d’Archæoptéryx, on trouve très tôt des oiseaux très différents (Confuciusornis, Enantiornithes, etc.). Cela indique une explosion soudaine d’oiseaux variés, et non une lente progression évolutive. Confuciusornis apporte une preuve supplémentaire que les oiseaux apparaissent d’emblée complets, avec plumes modernes et structures optimisées.
Il rend la narratif évolutif contradictoire car même si on interprète les structures filamenteuses de Sinosauropteryx comme des « proto-plumes », comment se fait il qu’il y avait déjà des plumes modernes totalement fonctionnelles (chez Confuciusornis)?
Il est difficile de comprendre qu’un oiseau avancé avec un bec comme Confuciusornis ait pu apparaître 10 millions d’années avant qu’il n’existe la moindre trace de ses ancêtres dinosaures à plumes.
L’un des principaux arguments de l’évolution est que l’ordre évolutif correspondrait supposément à la séquence fossile. Par conséquent, ce décalage flagrant avec les dinosaures-oiseaux représente un sérieux défi pour l’explication évolutionniste.
Anchiornis
Anchiornis huxleyi est un petit fossile (34 cm, 110 g) découvert en Chine, classé parmi les troodontidés (dinosaures théropodes carnivores). Il est présenté comme un « dino à quatre ailes » avec des plumes sur les bras mais aussi sur les jambes et même les pieds. Il est daté du Jurassique supérieur (161–151 Ma), donc plus ancien qu’Archæoptéryx (150 Ma). Il devait résoudre le « paradoxe temporel » car jusque-là, tous les prétendus dinosaures à plumes étaient plus jeunes que les vrais oiseaux.
Contrairement aux plumes asymétriques nécessaires au vol, celles d’Anchiornis sont symétriques et arrondies, donc non adaptées au vol battu.
Ses longues plumes sur les jambes et les pieds gêneraient même la course ce qui est en contradiction avec la théorie « cursoriale » (que les oiseaux descendraient de coureurs). Elles ne sont pas efficaces non plus pour un vrai vol plané ce qui est en contradiction avec la théorie arboricole (que les oiseaux descendraient de planeurs). En gros, Anchiornis ne correspond à aucune trajectoire évolutive claire.
Comme les dinosaures, Anchiornis ne possédait pas le système de sacs aériens aviaires, nécessaire au vol. Cela reste une barrière majeure au scénario dinosaure-oiseau.
Certes, Anchiornis est plus vieux qu’Archæoptéryx mais cela ne résout rien, car il est 30 Ma plus vieux que les autres troodontidés (famille de dinosaures théropodes) plus « dinoïdes ». Les évolutionnistes sont alors obligés de postuler un « grand vide fossile », autrement dit, tout un pan d’évolution hypothétique sans fossiles. La majorité des théropodes censés être ses ancêtres apparaissent bien après Archæoptéryx (au Crétacé).
Anchiornis accentue les contradictions dans la chronologie évolutive (les oiseaux comme Confuciusornis ou Archæoptéryx restent plus « avancés » mais apparaissent au même moment). La meilleure lecture est qu’Anchiornis représente une créature distincte, avec son propre design, et non un chaînon entre dinosaures et oiseaux.
Anchiornis, loin de résoudre le paradoxe de l’évolution des oiseaux, en crée de nouveaux. Il ne correspond ni à un coureur ni à un planeur, et ne rapproche pas les dinosaures du vol battu. Au contraire, il illustre la diversité d’animaux à plumes du registre fossile (peut-être un oiseau non volant), sans lien évolutif direct.
Ichtyornis et Hesperornis
Wang et al. (2017), PNAS est une analyse de la perte évolutive des dents chez les théropodes menant aux oiseaux, interprétée comme un cas de « troncature hétérochronique » dans le développement embryonnaire. La perte de dents chez les oiseaux serait plutôt une perte d’information (mutation, troncature du développement), ce qui ne démontre pas l’apparition d’une nouveauté évolutive. La régression est l’inverse de la macroévolution.
Certains évolutionnistes indiquent que ces deux espèces, retrouvées dans les strates du Crétacé, montre des mâchoires partiellement dentées et partiellement recouvertes d’un bec.
Ichthyornis et Hesperornis n’ont pas des becs et des dents de transition. Leurs dents étaient parfaitement adaptées à attraper du poisson. Leur bec corné à l’avant servait probablement de pince fine, ce qui était utile pour la préhension. Ce n’était pas une mâchoire « en train de devenir un bec », mais une structure stable et optimisée.
Si on suit strictement Darwin, on s’attend à une progression graduelle: reptiles -> mâchoire dentée -> dents qui se réduisent -> disparition -> puis bec. Cependant Ichthyornis (99.6 Ma à 70.6 Ma) et Hesperornis (100.5 Ma à 66 Ma) apparaissent après des oiseaux à bec complet (comme Confuciusornis, 130 à 113 Ma). Autrement dit, la « transition » attendue n’est pas pertinente, on a des mosaïques fonctionnelles et une diversité plus grande qu’aujourd’hui.
Microraptor
Xu et al. (2003) ont fait une découverte en Chine de fossiles d’une créature à quatre ailes (Microraptor), qui sont interprétés comme des formes transitoires du vol plané vers le vol battu des oiseaux. Microraptor est considéré comme une créature unique et complète, probablement un oiseau éteint à plumes. Il existe une controverse sur certains fossiles chinois, notamment sur Microraptor.13
Le Dr Storrs Olson, conservateur des oiseaux au Musée national d’histoire naturelle de la Smithsonian Institution à Washington D.C., a également noté que la fiabilité de Nature sur ce sujet est très douteuse:14
« L’idée des dinosaures à plumes et de l’origine des oiseaux à partir des théropodes est activement promue par un groupe de scientifiques zélés agissant de concert avec certains éditeurs de Nature et de National Geographic, qui sont eux-mêmes devenus des prosélytes déclarés et fortement biaisés de cette croyance. »
Le Dr Jonathan Sarfati conlut sur Microraptor:
« Bien que cette créature à quatre ailes ait probablement fait vendre beaucoup de journaux, de nombreuses questions subsistent quant à son statut d’ancêtre des oiseaux. Parmi elles :
- Le caractère douteux de la preuve elle-même, puisque tout provient de la même région que la fraude de l’Archaeoraptor, et que le premier spécimen nommé Microraptor en faisait en réalité partie.
- Les « datations » vont à l’encontre de ce que l’évolution prédirait, car M. gui est bien plus « jeune » que des oiseaux incontestés, y compris certains déjà dotés de becs.
- Cette dernière découverte réfuterait le paradigme dominant de la théorie cursoriale (origine du vol à partir de la course).
- Les transitions imaginées de l’animal terrestre au parachutiste, puis au planeur et enfin au voilier actif, auraient chacune nécessité l’apparition d’une quantité substantielle de nouvelles informations génétiques. De plus, les pattes en forme de colonnes d’un dinosaure, placées sous le corps, n’auraient pas pu s’écarter pour adopter une posture de vol plané sans modifications considérables de la hanche et des articulations.«
Prudence sur les fossiles brandis comme des chaînons manquants
Il faut être prudent quand on entend parler de « chaînon manquant ». On doit avoir à l’esprit des faux fossiles en Chine (cf. Archaeoraptor) et les fraudes connues sur l’évolution humaine (L’homme de Pilt Down, l’homme de Nebraska, les faux embryons d’Haeckel). Alan Feduccia (paléontologue évolutionniste critique du scénario théropode-oiseau) parle d’un « marché aux faux fossiles ».
« Archaeoraptor n’est que la partie émergée de l’iceberg. Il existe des dizaines de faux fossiles, et ils ont jeté une ombre inquiétante sur tout ce domaine. Lorsque vous allez dans ces expositions de fossiles, il est difficile de dire lesquels sont authentiques et lesquels sont faux. J’ai entendu dire qu’il existe une véritable usine de faux fossiles dans le nord-est de la Chine, dans la province du Liaoning, près des gisements où beaucoup de ces prétendus dinosaures à plumes ont été découverts.
Les revues comme Nature n’exigent pas que les spécimens soient authentifiés, et les spécimens repartent immédiatement en Chine, de sorte que personne ne peut les examiner. Ce sont peut-être des découvertes miraculeuses, peut-être des chaînons manquants comme on le prétend, mais il n’existe aucun moyen d’authentifier tout cela. »
Discover Dialogue: Ornithologist and Evolutionary Biologist Alan Feduccia Plucking Apart the Dino-Birds, Discover 24(2), Février 2003.
Le plus ancien oiseau du registre fossile
Protoavis est l’intrus qui dérange. Il a été découvert par Sankar Chatterjee en 1983 (Texas, Trias -225 Ma) donc 75 millions d’années plus ancien qu’Archæoptéryx. Il a une anatomie très avancée avec sternum en quille (comme les oiseaux modernes, indispensable au vol battu), une ceinture scapulaire avec système de poulie mais aussi des dents et une queue osseuse.
C’est un problème énorme (encore un) car il est plus ancien qu’Archæoptéryx, et même plus « avancé » morphologiquement, cela inverse complètement la chronologie évolutive.
Pourquoi les évolutionnistes rejettent Protoavis? Pas pour des raisons anatomiques car il ressemble bien à un oiseau mais parce qu’il n’entre pas dans le scénario « théropodes – oiseaux ». Du coup, beaucoup préfèrent l’ignorer ou le déclarer « controversé ».
Protoavis, s’il est bien interprété, ruine la chronologie évolutive, c’est un oiseau « moderne » bien avant ses supposés ancêtres dinosaures. La logique évolutionniste devient circulaire: on rejette ou on passe sous silence les fossiles qui n’entrent pas dans le schéma.
Protoavis présente le problème temporel récurrent du petit-fils qui apparaît avant le grand-père.
D’autres fossiles posent également problème:
« Le réexamen d’un fossile de la taille d’un moineau [Scansoriopteryx -164–159 Ma] provenant de Chine a conduit des chercheurs à remettre en question la croyance largement répandue selon laquelle les oiseaux ont évolué à partir de dinosaures terrestres qui auraient acquis la capacité de voler. »
— Birdlike fossil challenges notion that birds evolved from ground-dwelling dinosaurs — ScienceDaily, 9 juillet 2014
L’évolutionniste, Alan Feduccia, déclare:
« Au lieu de considérer que les oiseaux dérivent des dinosaures, Scansoriopteryx rétablit la validité de les voir comme une classe distincte, proprement aviaire et non dinosaurienne. »
Des restes de protéines dans le fossile original d’Archaeopteryx
Les chercheurs ont utilisé l’imagerie synchrotron pour analyser en profondeur un fossile d’Archaeopteryx (Bergmann et al., 2010). Ils ont identifié la chimie intacte de certaines parties, notamment des résidus de kératine et des traces de mélanosomes (structures qui portent les pigments des plumes).
Cela veut dire que non seulement la forme des plumes est conservée, mais qu’il y a des restes chimiques de protéines et pigments. Or, la kératine et les protéines en général sont considérées comme hautement instables sur de longues périodes. Même dans les meilleures conditions, elles devraient s’hydrolyser ou s’oxyder bien avant quelques millions d’années.
Archæoptéryx est daté à environ 153 millions d’années (Jurassique supérieur). Les modèles classiques de dégradation chimique disent que des protéines ne peuvent pas survivre plus que quelques milliers à quelques centaines de milliers d’années, même dans des conditions optimales (absence d’eau, basse température, etc.).
Donc retrouver des résidus de kératine et de pigments après 150 Ma est hautement improbable dans le cadre naturaliste. Cela contredit l’échelle de temps évolutive et semble plutôt indiquer une fossilisation beaucoup plus récente, compatible avec un modèle catastrophiste (comme celui du Déluge global).
Ces découvertes s’inscrivent dans une tendance observée depuis les années 1990/2000: plus on étudie les fossiles avec des outils modernes, plus on trouve de biomolécules en principe impossibles à préserver pendant des millions d’années. Cela soutient l’idée que ces fossiles sont jeunes, cohérents avec une fossilisation rapide durant le Déluge biblique (il y a quelques milliers d’années).
L’étude de Pan et al. 2019 (PNAS, vol. 116, n° 8, pp. 3018-3023) fait partie des découvertes les plus frappantes qui renforcent l’argument créationniste. Les chercheurs ont étudié plusieurs fossiles de dinosaures et d’oiseaux (dont Anchiornis) du Crétacé inférieur de Chine (gisement de Jehol, environ 130 millions d’années selon la chronologie évolutive).
En utilisant des techniques de pointe (spectroscopie infrarouge, spectroscopie Raman, etc.), ils ont trouvé des restes de kératine et de mélanosomes préservés dans les plumes fossilisées. Les mélanosomes (organites contenant les pigments) montraient même des signatures chimiques permettant d’inférer la couleur originelle des plumes (par ex. du noir ou de l’irisé).
Il ne s’agit donc pas uniquement d' »empreintes » ou de « moules », mais bien des résidus organiques détectables après 130 millions d’années supposés.
La kératine est une protéine fibreuse très fragile. En conditions naturelles, elle se dégrade en quelques milliers d’années maximum. Les pigments (mélanine) sont un peu plus stables, mais certainement pas sur 100+ millions d’années. Pan et al. reconnaissent que ces résultats montrent une préservation moléculaire directe donc pas seulement des « minéralisations » mais des molécules originales. Cela contredit totalement l’idée d’une fossilisation sur des dizaines de millions d’années.
Les auteurs invoquent une préservation exceptionnelle dans les sédiments de Jehol, grâce à des conditions chimiques particulières (faible oxygène, minéralisation rapide) mais ces explications restent ad hoc, car on retrouve des molécules organiques similaires dans des fossiles très variés et de nombreux contextes géologiques (dinosaures, reptiles marins, mammifères, poissons).
En revenant sur le cas de Microraptor, le Dr Angela Milner, conservatrice associée de paléontologie au Muséum d’Histoire naturelle de Londres, avait été citée comme suit:15
« Avec les dino-oiseaux de Liaoning, la matière elle-même, la kératine des plumes, est toujours présente. La découverte est stupéfiante. »
Le chimiste créationniste Jonathan Sarfati répond:16
« …la kératine est une protéine, c’est-à-dire un type de polymère de condensation, et elle est sujette à la dégradation en ses monomères. Les preuves bien connues de la chimie observationnelle montrent que les protéines ne peuvent absolument pas persister durant des millions d’années. »
Les traces d’oiseaux avant Archæoptéryx
L’étude menée par Abrahams et Bordy en 2023, intitulée « The oldest fossil bird-like footprints from the upper Triassic of southern Africa » et publiée dans PLoS ONE, révèle la découverte de traces fossiles ressemblant à celles d’oiseaux dans des strates datant du Trias supérieur (environ 230-201 millions d’années), en Afrique australe.
Les chercheurs ont découvert des empreintes fossiles qui ressemblent à celles d’oiseaux dans des strates géologiques datées du Trias supérieur, soit il y a environ 212 millions d’années. Ces empreintes ont été trouvées dans des formations rocheuses de l’Afrique du Sud.
Les empreintes présentent des caractéristiques similaires à celles des oiseaux modernes, comme des doigts fins et allongés. Cependant, elles proviennent d’une époque bien antérieure à celle des premiers oiseaux officiellement connus, tels qu’Archæoptéryx, qui est daté du Jurassique supérieur (environ 150 millions d’années dans le référentiel évolutif).
Les auteurs de l’étude discutent de la possibilité que ces empreintes aient été laissées par des dinosaures théropodes primitifs ou des archosaures capables de produire des traces ressemblant à celles des oiseaux.
Cette découverte remet en question une fois de plus certaines hypothèses sur l’évolution et la chronologie de l’apparition des oiseaux. Si des empreintes ressemblant à celles des oiseaux existent dans le Trias supérieur, cela pourrait indiquer que des traits ressemblant à ceux des oiseaux, tels que la morphologie des pieds et une démarche bipède, existaient bien avant l’apparition des oiseaux reconnus dans le registre fossile.
Sans l’influence évolutionniste, il devient tout à fait facile d’accepter qu’il s’agit de traces d’oiseaux. Si « la créature marchait comme un oiseau », alors c’était probablement un oiseau! Cela n’est pas très étonnant car cette caractéristiques de traces de pas qui apparait avant les créatures qui les ont formées est courante dans le registre fossile, cela s’explique bien dans le déluge mais très difficilement par l’évolution:
Le Dr Jake Hebert fait bien le point sur les diverses traces retrouvées d’oiseau et la réaction évolutionniste. Il rapporte par exemple que le paléontologue amateur et évolutionniste Jerry MacDonald avait mené des recherches paléontologiques novatrices au Nouveau-Mexique. Parmi ses découvertes, il avait trouvé des empreintes dans des roches du Jurassique qui ressemblaient étrangement à des traces d’ours, d’oiseaux et de singes.
« Les empreintes fossiles que MacDonald a collectées incluent un certain nombre de ce que les paléontologues aiment appeler des « problematica ».
Sur une piste, par exemple, une créature à trois doigts a apparemment fait quelques pas, puis a disparu — comme si elle s’était envolée. « Nous ne connaissons aucun animal à trois doigts au Permien [298.9 Ma – 251.902 Ma selon le référentiel évolutif] », souligne MacDonald. « Et il n’est pas censé y avoir d’oiseaux. »
Il possède plusieurs pistes où des créatures semblent marcher sur leurs pattes arrière, d’autres qui paraissent presque simiennes [semblables à des singes].
Sur une paire de tablettes de siltstone, je remarque des empreintes inhabituellement grandes, profondes et inquiétantes, chacune avec cinq marques de doigts arqués, comme des griffes. Je fais remarquer qu’elles ressemblent exactement à des traces d’ours. « Oui », dit MacDonald à contrecœur, « c’est bien ce qu’elles semblent être. » Les scientifiques s’accordent à dire que les mammifères ont évolué bien après la période permienne, et pourtant ces empreintes sont clairement permiennes.«
Stewart, D. 1992. Petrified Footprints: A Puzzling Parade of Permian Beasts. Smithsonian. 23 (4): 70–79.
En tenant compte de ses diverses traces d’oiseau (voir article d’Hebert), probablement laissées lorsque des sédiments mous étaient brièvement exposées durant le déluge, un esprit frais et neutre perçoit aisément que la science ne valide pas le scénario de l’évolution de théropode à oiseau.
Des évolutionnistes dissidents?
Il y a une logique d’apparition soudaine et de stase dans le registre fossile. Cela cadre bien plus naturellement avec le modèle créationniste qu’évolutionniste.
« L’évolution semble s’être produite par à-coups — du moins, c’est ce que montrent les archives fossiles. Des trilobites aux ptérodactyles, des ammonites à l’Archaeopteryx, les scientifiques observent le même schéma : de brèves flambées d’innovation au cours desquelles une seule espèce, ou une branche de l’arbre de la vie, donne naissance à une grappe de nouvelles ramifications, suivies de longues périodes de stabilité dominées par le même statu quo. »
H. Powell, « Forest songsters evolved in an early burst of innovation », communiqué de presse de l’Université Cornell, 9 juillet 2008, rapportant une recherche publiée dans: D. L. Rabosky et I. J. Lovette, 2008, Density-dependent diversification in North American wood warblers, Proceedings of the Royal Society B, 275(1649), pp. 2363–2371.
Le problème évolutionniste est de vouloir voir du temps long dans le registre fossile. Si on raisonne avec le modèle de zonage écologique diluvialiste, on comprend que ces couches se sont déposées rapidement et que les étapes géologiques sont en fait des étapes du déluge qui a duré un an.
Le Dr Alan Feduccia, une autorité mondiale sur les oiseaux à l’Université de Caroline du Nord à Chapel Hill et un évolutionniste lui-même (voir Feduccia vs Creationists), dit:
«Les paléontologues ont essayé de transformer l’archéoptéryx en un dinosaure terrestre à plumes. Mais ce n’en est pas un. C’est un oiseau, un oiseau perché. Et aucune quantité de «paléobalbutiement» ne changera cela».
Feduccia, A.; cité dans: V. Morell, Archaeopteryx: Early Bird Catches a Can of Worms, Science 259(5096):764–65, 5 February 1993.
Il existe un certain nombre d’évolutionnistes en désaccord avec l’idée de l’évolution des théropodes aux oiseaux via l’Archæoptéryx:
- Alan Feduccia,
- Larry Martin,
- Theagarten Lingham-Soliar,
- John Ruben,
- Storrs Olson.
Et parmi ceux qui y croient il y a parfois des admissions parlantes:
« L’hypothèse d’un ancêtre bipède, bien que fortement soutenue par la logique, possède peu de preuves directes dans les archives fossiles pour l’appuyer ou la réfuter. »
Shipman, P., 1998, Taking Wing: Archaeopteryx and the Evolution of Bird Flight, p. 84, Simon and Schuster, New York.
L’origine des dinosaures: un conundrum similaire comme avec l’origine des oiseaux?
« Désormais, l’attention est passée de la façon dont ils ont disparu à la question de leur origine. »
« Les dinosaures ont coexisté avec leurs ancêtres reptiliens, plutôt que de les remplacer. »
« A new fossil find in Brazil rewrites the history of the dinosaurs. » The Economist, 10 novembre 2016.
« Cependant, la manière et le moment de l’origine et de la diversification des dinosaures restent, à ce jour, non résolus. »
— Bernardi et al., 2018, Nature Communications.
« D’après ma lecture du registre fossile des dinosaures, aucun ancêtre direct n’a été découvert pour aucune espèce de dinosaure. Hélas, ma liste d’ancêtres dinosauriens est vide. »
Dr David Weishampel, Université Johns Hopkins, auteur principal de The Dinosauria – e-mail communication with Dr. Carl Werner. Cité dans Werner, C. 2007. Evolution: The Grand Experiment. Green Forest, Arkansas: New Leaf Press, 126.
« Nous étudiions les empreintes dans les Dolomites [séquences rocheuses du nord de l’Italie] depuis un certain temps, et il est étonnant de constater à quel point le passage de « aucun dinosaures » à « tous les dinosaures » a été net. »
Dinosaurs ended—and originated—with a bang! Science Daily. Publié sur sciencedaily.com le 16 Avril 2018.
Conclusion
Dans le modèle évolutif néo-darwinien, on s’attend à ce que la diversité (nombre d’espèces) et la disparité (écart morphologique, variété de plans corporels) augmentent progressivement au cours du temps, avec de petites différences au début puis de plus en plus grandes.
Ce que l’on observe toutefois dans le registre fossile, notamment lors de l’explosion cambrienne, c’est l’inverse: la disparité est maximale dès le début (apparition brutale de la plupart des grands plans corporels animaux). Ensuite, la diversité spécifique (nombre d’espèces par plan corporel) augmente, mais dans les limites des « plans » déjà présents. Ce verger de la vie (suspendu diraient certains créationnistes), est l’inverse de l’arbre de la vie de Darwin et d’Haeckel, et c’est pourtant ce qu’on observe dans le registre.
- Si Archæoptéryx est un oiseau qui pouvait voler de manière performante, comme les oiseaux modernes, qu’est-ce qui le rend « transitionnel »?
- Il présente des caractéristiques mosaïques, mais pourquoi cela signifierait obligatoirement qu’il est une transition? Qu’est-ce qu’il y a de plus fort et convaincant que chez la chauve-souris et l’ornithorynque?
- Archæoptéryx a donc des plumes parfaitement adaptées au vol battu, il n’est donc pas une transition à cet égard, mais tout aussi optimisé qu’un oiseau « moderne ».
John Ruben a démontré que les poumons des théropodes n’auraient pas pu évoluer vers ceux des oiseaux. On a donc un système respiratoire optimisé pour le vol et des plumes parfaites pour le vol. Qu’y a-t-il de transitionnel chez Archæoptéryx au final? Seulement ses griffes, ses dents et sa queue?
D’autres oiseaux fossiles (Hesperornis, Ichthyornis) avaient aussi des dents. C’est juste une variation. La queue osseuse longue? mais elle est plumée et aérodynamique, ce qui en fait une queue d’oiseau et non de reptile.
Des griffes aux ailes? elles sont présentes aussi chez certains oiseaux modernes comme le hoatzin et le touraco chez les jeunes, elles sont utilisées pour grimper (les ptérosaures avaient aussi des doigts griffus). C’est un outil supplémentaire pour grimper chez certains oiseaux.
Chaque aile d’Archæoptéryx possédait trois doigts distincts, terminés par des griffes fonctionnelles. Ces doigts étaient libres et griffus, contrairement aux oiseaux modernes où les doigts sont fusionnés dans l’aile. Cela suggère qu’Archaeopteryx pouvait s’agripper aux branches, grimper ou s’aider de ses ailes pour manœuvrer dans la végétation.
Darwin attendait des séries abondantes et continues de formes intermédiaires montrant des transformations graduelles entre grands groupes.
Paradoxalement pour le modèle évolutionniste, Archæoptéryx avait plus d’outils que les oiseaux modernes. On ne passe pas du simple au complexe. Archaeopteryx avait:
- des griffes aux ailes utiles pour grimper, s’agripper aux branches, manœuvrer dans la végétation.
- des petites dents permettaient sans doute une alimentation plus variée (insectes, petits animaux, peut-être même fruits durs).
- une queue longue et osseuse pouvait servir de gouvernail en vol ou en plané, un stabilisateur naturel.
Résume sur les transformations nécessaires pour passer de Théropode à Oiseau
| Domaine | État chez les théropodes | État chez les oiseaux | Fonction nouvelle requise |
|---|---|---|---|
| Locomotion / Bassin | Fémur mobile, sert à courir (comme humains) | Fémur fixé contre le corps, rôle clé dans la respiration | Permet respiration par sacs aériens et vol |
| Respiration | Système piston hépatique (comme crocodiles) | Système de sacs aériens + flux d’air unidirectionnel | Haut rendement énergétique pour vol |
| Sternum | Plat ou réduit | Carène (bréchet) développé | Point d’ancrage des muscles de vol |
| Clavicules | Absentes ou séparées | Fusion en furcula (fourchette) | Ressort élastique durant le battement d’ailes |
| Queue | Longue queue osseuse lourde | Pygostyle court et rigide | Support des plumes caudales pour stabilité en vol |
| Bras vers Ailes | Bras courts, doigts griffus | Ailes longues, os fusionnés (carpometacarpe) | Portance + contrôle du vol |
| Doigts | Trois griffes manuelles fonctionnelles | Réduction/fusion, parfois griffes (ex : hoazin juvénile) | Support des rémiges primaires |
| Plumes | Absentes (écailles kératiniques) | Plumes complexes, asymétriques, imbriquées | Aérodynamique, isolation thermique |
| Vision | Vision correcte mais limitée | Vision diurne ultra-performante, fovéa | Pilotage en vol, détection proies |
| Oreille interne | Moins développée pour équilibre terrestre | Développée (canaux semi-circulaires) | Orientation 3D en vol |
| Cerveau | Petits lobes optiques et cérébelleux | Hypertrophie cervelet + vision | Coordination fine du vol |
| Métabolisme | Ectotherme / mésotherme (débattu) | Endotherme strict (sang chaud) | Vol exige très haut métabolisme |
| Dentition | Dents coniques ou tranchantes | Perte des dents, apparition du bec | Allègement du crâne, spécialisation alimentaire |
| Appareil digestif | Estomac simple | Bec + gésier + cailloux | Mastication interne sans dents |
| Reproduction | Œufs calcifiés, plusieurs ovaires | Un seul ovaire fonctionnel | Allègement du corps pour vol |
| Développement embryonnaire | Placodes d’écailles | Placodes de plumes + voies de signalisation (Shh, BMP, Wnt) | Différenciation plumes spécialisées |
| Circulation sanguine | Cœur 3 ou 4 cavités mais efficacité limitée | Cœur 4 cavités très efficace | Débit élevé pour endurance en vol |
Références:
- Johnson, J. S. 2021. Are Birds “Cousins” to Reptiles? Acts & Facts. 50 (12): 11.
- Feduccia, A., T. Lingham-Soliar, and J. R. Hinchliffe. 2005. Do Feathered Dinosaurs Exist? Testing the Hypothesis on Neontological and Paleontological Evidence. Journal of Morphology. 266 (2): 125-166.
- « Arracher le nez » est un jargon d’aviation pour dire faire la rotation: on cabre l’appareil pour lever le nez (augmenter l’angle d’attaque) à la vitesse de rotation afin de quitter le sol. Avec un avion, tirer sur le manche permet une profondeur vers le haut et plus de portance et de moment cabreur, le nez se lève, la roulette avant quitte la piste. Chez l’oiseau, la queue déployée et abaissée crée un moment cabreur similaire, aidant à lever l’avant du corps et à garder le contrôle à très basse vitesse.
- https://www.sci.news/paleontology/archaeopteryx-feeding-structures-flight-14539.html.
- O’Connor, J. K. et al. 2026. Avian Features of Archaeopteryx Feeding Apparatus Reflect Elevated Demands of Flight. The Innovation. 7 (2).
- https://www.icr.org/article/archaeopteryx-myth-transitional-fossil/ voir les sources séculières dans son article.
- https://www.icr.org/article/dinosaurs-reptilian-metabolic-adaptation.
- https://www.icr.org/article/dinosaurs-designed-cold-blooded.
- Morell, V. 1996. A Cold, Hard Look at Dinosaurs. Discover. 17 (12): 98-108..
- Ruben, J. A. 1996. The Metabolic Status of Some Late Cretaceous Dinosaurs. Science. 273 (5279): 1204-1207.
- Erickson, G. M., et al. 2017. Dinosaur incubation periods directly determined from growth-line counts in embryonic teeth show reptilian-grade development. Proceedings of the National Academy of Sciences. 114 (3): 540-545. DOI: 10.1073/pnas.
- https://www.icr.org/article/dinosaur-eggs-not-bird-like-after-all.
- https://creation.com/new-four-winged-feathered-dinosaur.
- https://creation.com/sensationalistic-unsubstantiated-tabloid-journalism.
- Lee, A., Winged dinosaur fossils may show how life took to the skies, Straits Times, Singapore, 24 January 2003.
- https://creation.com/new-four-winged-feathered-dinosaur.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!