
Les Mammifères ont-ils évolué à partir des Thérapsides (Reptiles Mammaliens)?
Les synapsides non-mammaliens (SNM)1 sont généralement considérés, d’un point de vue évolutionniste, comme des formes transitionnelles entre des animaux ressemblant à des reptiles et les mammifères. Ils devraient selon cette hypothèse afficher de plus en plus de caractéristiques de mammifères au fil du registre fossile, surtout au sein de six sous-groupes therapsidiens.

S’y retrouver entre synapsides, thérapsides et reptiles mammaliens
Les synapsides
Les synapsides forment un grand groupe de vertébrés à part entière qui inclut à la fois les ancêtres des mammifères et les mammifères eux-mêmes. Ils se distinguent par la présence d’une seule ouverture (fenêtre temporale) de chaque côté du crâne, juste derrière l’orbite, où s’insèrent des muscles de la mâchoire. Cette caractéristique permet une meilleure attache des muscles et une force de morsure accrue.
Ils incluent les pélycosaures, les thérapsides et finalement les mammifères. Les premiers synapsides, comme les pélycosaures (dont Dimetrodon), auraient selon les évolutionnistes « évolué » durant le Carbonifère, il y a environ 320 millions d’années. Les synapsides ne sont pas des « reptiles » dans le sens strict, bien qu’on les associe parfois à tort à des reptiles primitifs en raison de leur apparence et de leur mode de vie similaire.
Les thérapsides
Les thérapsides forment un sous-groupe des synapsides, les évolutionnistes avancent qu’ils auraient évolué à partir de certains pélycosaures au cours du Permien, il y a environ 275 millions d’années. Parmi les thérapsides, les cynodontes seraient le groupe le plus proche des mammifères, ils sont donc considérés par les évolutionnistes comme leurs ancêtres directs.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
Les « Reptiles Mammaliens »
Le terme « reptiles mammaliens » est un terme obsolète et parfois trompeur utilisé autrefois pour désigner certains groupes de thérapsides, en particulier les cynodontes et d’autres thérapsides proches des mammifères.
Ce terme est apparu parce que les premiers paléontologues voyaient les thérapsides comme des « reptiles » qui présentaient des traits rappelant les mammifères. Cependant, il est aujourd’hui reconnu que les thérapsides et leurs descendants, les cynodontes, appartiennent à une lignée distincte de celle des reptiles véritables (sauropsides).
Aujourd’hui, on évite le terme « reptiles mammaliens », car il est trompeur : les thérapsides ne sont pas des reptiles, mais des synapsides, une lignée séparée. Le terme peut cependant encore être utilisé de manière informelle pour parler de certains thérapsides qui ressemblent à des mammifères, notamment dans les ouvrages anciens.
Une perspective créationniste
Emily Anderson et Matthew A. McLain proposent une perspective créationniste (selon laquelle Dieu a créé des « genres » d’animaux distincts). Ils ont réévalué cette interprétation de transition en étudiant la structure des mains et des pieds des SNM2.
Dans ce contexte, la formule digitale (nombre de phalanges par doigt) est un élément clé. Les amniotes, par exemple, ont classiquement une formule digitale reptilienne (2-3-4-5-3 pour la main, 2-3-4-5-4 pour le pied), tandis que les mammifères ont une formule plus homogène (2-3-3-3-3). Les auteurs ont constaté que dans les archives fossiles, divers synapsides non-mammaliens (SNM) montrent des formules digitales qui peuvent être reptiliennes, mammaliennes ou même uniques.
Alors que la recherche antérieure associait ces formules digitales aux relations phylogénétiques évolutionnaires des SNM, Anderson et McLain ont adopté une approche stratigraphique. En analysant la littérature et en regroupant les SNM en fonction de leur stratigraphie, ils ont observé que la plupart des groupes therapsidiens présentent des formules digitales proches de celles des mammifères dès leurs premières apparitions fossiles, sauf quelques exceptions. Cela signifie qu’il n’y a pas de progression évolutive évidente vers une formule digitale de type mammalien dans le temps.
Les résultats mettent en doute l’idée d’une transition évolutive progressive dans les doigts des SNM vers les mammifères et posent la question de l’origine de la grande variabilité dans les formules digitales au sein de ce groupe.
Pas d’évolution graduelle observable
Il n’y a pas de fossiles complets montrant une progression claire et continue entre reptiles mammaliens et mammifères modernes. L’une des critiques importantes est basée sur les différences anatomiques et physiologiques entre les reptiles mammaliens et les mammifères, en particulier les caractéristiques des mâchoires, des os de l’oreille, et des types de reproduction (oviparité chez beaucoup de reptiles contre viviparité chez la plupart des mammifères). Ces différences sont trop significatives pour être le produit de mutations progressives et d’une sélection naturelle.
Il y a aussi la difficulté pour des organes intermédiaires (comme la transformation des os de la mâchoire en os de l’oreille chez les mammifères) de fonctionner efficacement pendant la transition. De telles transformations rendraient temporairement l’organisme moins viable ce qui aurait dû empêcher sa survie selon la logique de la sélection naturelle.
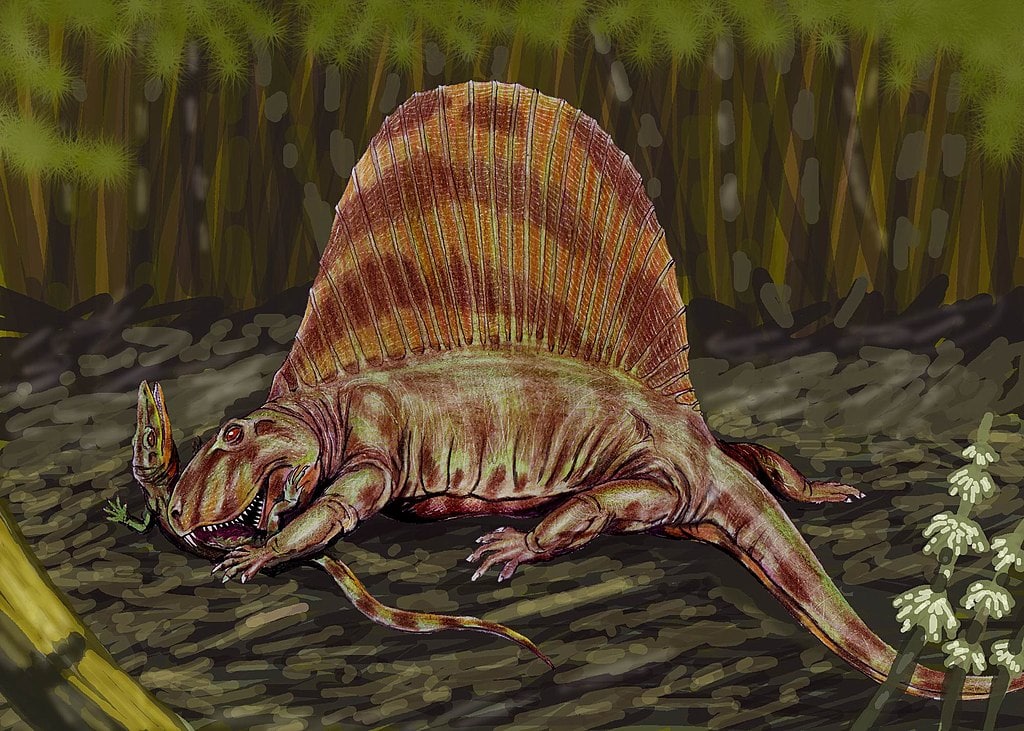
Les évolutionnistes voient dans les fossiles de thérapsides une transition graduelle de caractéristiques reptiliennes vers mammifères mais il faut être conscient des différences substantielles entre les pélycosaures anciens et les thérapsides plus récents. La paléontologie ne documente pas les transitions des pélycosaures vers les thérapsides. Dimetrodon apparait soudainement dans le registre fossile, complet et entièrement formé. Il est intéressant que les évolutionnistes voit en cet animal un lien possible avec les mammifères, nonobstant totalement « l’âge des dinosaures » et les liens avec d’autres reptiles.
Arbre de la vie VS Verger de la création
En fait les évolutionnistes observent eux mêmes les prédictions créationnistes. Le modèle biblique prévoit différents genres distincts au sein desquels un certain nombre de variation est possible. On appelle cela le verger de la création, une forêt de la vie bien plus qu’un arbre de la vie. Chaque arbre de la forêt de la vie est indépendant et va ensuite se brancher et varier dans une certaine limite.
Anderson et Sue ont noté que « l’arbre phylogénétique des mammifères du Mésozoïque, tel que le soutiennent les preuves actuelles, est un vaste buisson évolutif »3. Un « buisson » ne soutient pas l’idée d’un « arbre » et de la macro évolution mais plutôt le modèle créationniste.
L’apparition des poils chez les mammifères
La présence de poils chez les mammifères est également un problème de plus à expliquer:
« La présence de poils chez les mammifères peut sembler une question triviale, mais la chronologie exacte de l’apparition des poils chez les mammifères est extrêmement rare. L’origine développementale des poils est tout aussi mystérieuse ; à ce jour, aucun scénario bien testé (ou testable) n’a été proposé pour expliquer les transformations qui peuvent expliquer l’évolution des poils. »4

Les poils et les plumes se développent à partir de follicules complexes situés au plus profond de la couche dermique de la peau, contrairement aux écailles reptiliennes, leurs précurseurs présumés.
Les glandes mammaires
Le lait des glandes mammaires est encore plus révélateur de ce groupe que les poils. Bien qu’ils ne le disent pas souvent à haute voix, les évolutionnistes pensent que les mamelles ont évolué à partir des glandes sudoripares (absentes chez les reptiles) et le lait à partir de la sueur. Mais ils ne décrivent généralement pas ce qui est arrivé aux bébés mammifères pendant la longue et lente transition, étape par étape, d’une solution aqueuse fine et salée d’urée et de diverses toxines à un liquide épais et nutritif, riche en protéines, en sucre et en graisse, et même en anticorps qui combattent les maladies.

Morgacunodon
Les évolutionnistes pensent que les mammifères ont évolué à partir de synapsides, un groupe de vertébrés qui avait une mâchoire et des os de l’oreille différents de ceux des mammifères modernes. Selon la théorie de l’évolution, le processus de transition a impliqué des changements progressifs dans la mâchoire et l’oreille, avec des fossiles comme Morganucodon montrant une mâchoire double (une caractéristique supposée intermédiaire).

Pour les évolutionnistes Morganucodon représente la forme de transition la plus définitive entre les reptiles et les mammifères, il possède l’articulation de la mâchoire de type mammifère côte à côte avec l’articulation de la mâchoire de type reptile. Morgacunodon avait un seul os d’oreille et un complément complet d’os reptiliens dans la mâchoire inférieure. Mais une autre interprétation des preuves est que Morganucodon était un mammifère et non une espèce transitionnelle.
Les évolutionnistes disent que Morganucodon aurait eu une double articulation de la mâchoire : une articulation dentaire-squamosale (comme celle des mammifères) et une articulation quadrate-articulaire (comme celle des reptiles). Cependant, Marc Surtees note que les os de la mâchoire et de l’oreille retrouvés chez Morganucodon sont fragmentaires5, souvent absents ou désarticulés, et que cette double articulation est en partie déduite.
De nombreux aspects de la structure de la mâchoire et de l’oreille moyenne chez Morganucodon sont des reconstructions basées sur des inférences, et non sur des preuves fossiles complètes. Par exemple :
- La structure du squamosal (un os impliqué dans l’articulation de la mâchoire chez les mammifères) est en partie supposée, car l’orientation de cet os est incertaine.
- La présence du quadratojugal, un os supposé faire partie de l’articulation, est uniquement déduite, car cet os n’a pas été retrouvé.
Cela signifie que les interprétations reliant ces structures à une « mâchoire double » et une oreille intermédiaire sont en grande partie hypothétiques.
Morganucodon serait plutôt un mammifère complet, possédant une articulation de mâchoire dentaire-squamosale et une oreille moyenne typique des mammifères, avec des os de l’oreille (par exemple, le quadrate pourrait être interprété comme un incus, un os de l’oreille des mammifères modernes). Le petrosal (la région de l’os temporal entourant l’oreille interne) est décrit comme pleinement mammalien ce qui soutient cette conclusion.
Les preuves disponibles correspondent davantage à une mâchoire et une oreille de type mammalien, ce qui soutient l’idée que Morganucodon était déjà un mammifère.
L’absence de preuves fossiles pour l’évolution des mammifères
R. L Carrol déclare:
« La transition entre les pélycosaures et les thérapsides n’a pas été documentée. »6
Carroll déclare encore:
« Les ancêtres des mammifères [à la page suivante on comprend qu’il fait référence aux synapsides] sont identifiés à partir du même horizon et de la même localité que le premier reptile conventionnel, Hylonomus, dans le Pennsylvanien ancien de Joggins, en Nouvelle-Écosse. » (Carroll, 361.) »
La vérité est que lorsque les thérapsides apparaissent pour la première fois, ils diffèrent considérablement des pélycosaures et il n’existe aucune espèce intermédiaire reliant de manière plausible une espèce connue des deux ordres. L’affirmation selon laquelle les thérapsides descendent des pélycosaures repose sur l’hypothèse de l’évolution et sur la croyance que parmi les créatures connues pour avoir précédé les thérapsides dans les archives fossiles, les pélycosaures sont la source la plus envisageable de l’espèce ancestrale.
Les évolutionnistes essaient de relier des points ensembles et ils doivent trouver des candidats probables. Certains évolutionnistes sont obligés de faire appel à « l’évolution convergente », c’est à dire qu’il y aurait eu des « évolutions indépendantes » mais cela trahit l’idée de « l’arbre de la vie » et renforce le « verger de la création ».
Les cynodontes
Des thérapsides, l’évolution aurait poursuivi son chemin vers les Cynodontes, qui auraient à leur tour évolué en mammifères primitifs. Là encore les fossiles intermédiaires sont manquants pour étayer cette évolution. L’arborescence évolutive est un peu comme suit:
- Synapsides : C’est le groupe principal qui inclurait tous les animaux à l’origine des mammifères, avec une ouverture temporale unique dans le crâne.
- Pélycosaures : ils seraient des synapsides primitifs, parmi les premiers à montrer une fenêtre temporale.
- Thérapsides : Ce serait le sous-groupe de synapsides « évolué », avec des traits plus proches des mammifères (position des membres, dentition complexe).
- Cynodontes : Ce serait le groupe de thérapsides particulièrement proche des mammifères, ils possèdent des caractéristiques très similaires aux mammifères modernes.
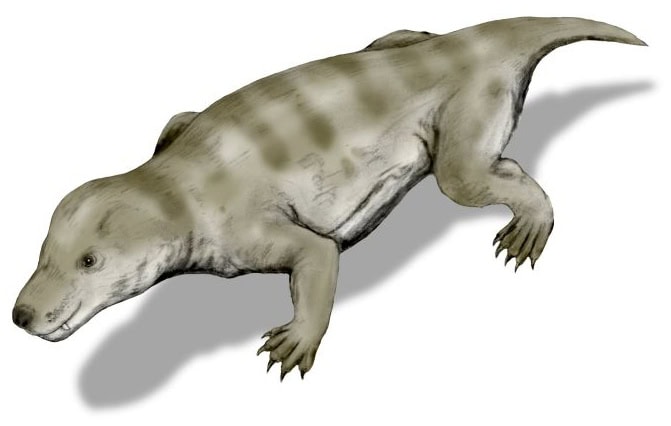
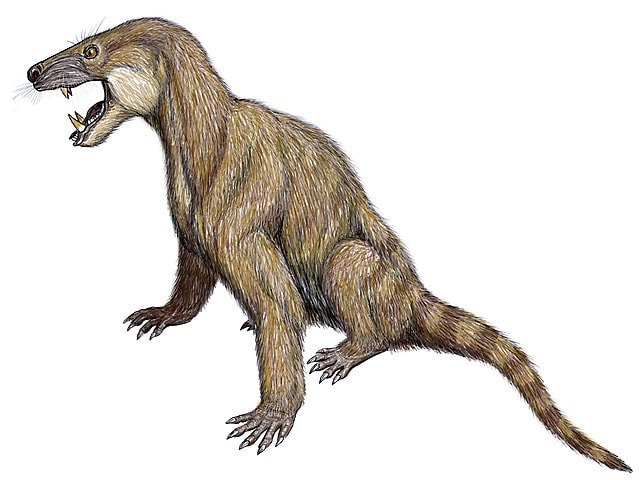
On impute généralement cette incapacité à retracer la transition du cynodonte au mammifère à la rareté des fossiles. Carroll écrit :
« Malheureusement, les archives des ancêtres immédiats des mammifères deviennent moins complètes au cours du Trias supérieur » (Carroll, 392).
Il existe cependant des fossiles d’au moins deux superfamilles, trois familles et sept genres de cynodontes « avancés » du Trias supérieur (Carroll, 624). Il se trouve qu’aucun d’entre eux ne convient comme transition vers les mammifères.
George Gaylord Simpson présente la question sous un angle quelque peu différent :
L’événement le plus déroutant de l’histoire de la vie sur terre est le passage du Mésozoïque, l’ère des reptiles, à l’ère des mammifères. C’est comme si le rideau se baissait soudainement sur la scène où tous les rôles principaux étaient tenus par des reptiles, en particulier des dinosaures, en grand nombre et d’une variété déconcertante, et se levait aussitôt pour révéler le même décor mais une distribution entièrement nouvelle, une distribution dans laquelle les dinosaures n’apparaissent pas du tout, les autres reptiles sont des figurants, et tous les rôles principaux sont tenus par des mammifères de toutes sortes à peine évoqués dans les actes précédents. (Kerwin, 42.)
Kerwin, Carlotta and others (editors). 1972. Life Before Man. Time-Life Books. New York.
Kemp a déclaré:
L’apparition soudaine de nouveaux taxons supérieurs, de nouvelles familles et même de nouveaux ordres, immédiatement après une extinction massive, avec toutes les caractéristiques plus ou moins développées, implique une évolution très rapide. . . . Il est possible qu’il s’agisse d’un artefact et que les nouveaux taxons aient eu une longue histoire avant d’apparaître dans les archives fossiles, au cours de laquelle ils ont progressivement acquis leurs caractéristiques. Cependant, dans aucun cas une histoire aussi longue n’est connue, même par un seul spécimen, et il est donc beaucoup plus raisonnable d’accepter que des taux très élevés d’évolution morphologique se produisent de manière caractéristique après une extinction massive (Kemp, 327).
Kemp, T. S. 1982. Mammal-like Reptiles and the Origin of Mammals. Academic Press. New York.
Conclusion
La récurrence des « explosions » de la vie dans le registre fossile s’explique par la création de la Genèse, associée au déluge biblique de Noé, un événement progressif rempli de mini-événements qui ont duré un an. Les groupes d’organismes (le rang taxonomique genre ou famille) ont été créés au cours de la semaine de la création.
Lors du déluge, des biomes ont été enfouis l’un après l’autre, selon les montées et retraits des eaux. Ces montées et retraits des eaux ont à voir avec des tsunamis de très grandes ampleurs7 mais aussi par l’élévation du nouveau sol océanique chaud, étant plus haut que l’ancien sol océanique froid.
Les groupes d’organismes emportés dans les différentes strates reflètent l’ordre général dans lequel ils ont été pris par les eaux. Le déluge a commencé dans les océans puis s’est élevé jusqu’à recouvrir toutes les montagnes antédiluviennes. D’autres montagnes (comme les Alpes, l’Himalaya) ont surgi à la fin et après le déluge, emportant avec eux des fossiles marins dans leurs sommets.
A la suite du déluge, les organismes se sont répandus à travers le monde et se sont adaptés à différentes niches écologiques (spéciation et adaptation au sein d’un genre).
Découvrez plein d’articles sur le Déluge de Noé dans la bibliothèque du site:
Références:
- Les termes « synapside », « thérapside » et « reptiles mammaliens » sont souvent utilisés pour décrire des groupes d’animaux dans la lignée évolutive menant aux mammifères, mais ils représentent des catégories différentes dans cette transition.
- Anderson, Emily and McLain, Matthew A. (2023) « Does the Fossil Record of Non-Mammalian Synapsid Digits Show an Increasing « Mammal-ness »?, » Proceedings of the International Conference on Creationism: Vol. 9, Article 56. DOI: 10.15385/jpicc.2023.9.1.49.
- Anderson and Sues, 2007, 341. Voir aussi Y.I Wolf, I. B. Rogozin, N. V. Grishin and E. V. Koonin, 2003, Genome trees and the tree of life, Trends in Genetics, 18 (9): 472-479.
- Anderson and Sues, 2007, 153.
- https://creation.com/morganucodon.
- R. L Carroll, 1997, Patterns and Processes ov Vertebrate Evolution. Cambridge, UK: Cambridge University Press, 397.
- Baumgardner, John and Navarro, Evan (2023) « The Role of Large Tsunamis in the Formation of the Flood Sediment Record, » Proceedings of the International Conference on Creationism: Vol. 9, Article 13. DOI: 10.15385/jpicc.2023.9.1.22.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!