

L’Évolution des Baleines est-elle démontrée?
Les évolutionnistes présentent une transition progressive des mammifères terrestres vers les baleines aquatiques à travers une série de formes fossiles:
- Pakicetus (semi-terrestre),
- Ambulocetus (comme une loutre géante),
- Rodhocetus, Basilosaurus, jusqu’aux cétacés modernes.
Ce scénario est souvent cité comme une preuve très forte de la macroévolution. Mais l’interprétation des fossiles est très spéculative. Les créationnistes insistent sur le nombre limité de fossiles et leur caractère fragmentaire. Par exemple Pakicetus a été initialement reconstruit comme aquatique, mais les fossiles complets ultérieurs montrent qu’il était entièrement terrestre, avec des membres adaptés à la marche. Il faut faire attention aux reconstructions artistiques trompeuses faites à partir de quelques fragments crâniens.
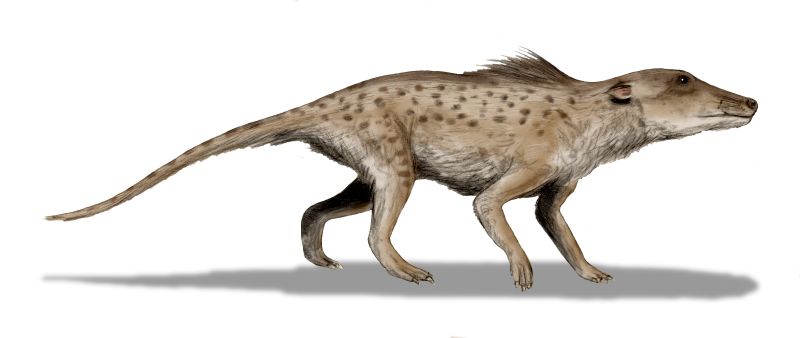
Voici comment les évolutionnistes l’avaient imaginé au début:
Les structures anatomiques
Les structures anatomiques des cétacés modernes (nageoires, souffles, oreilles spécialisées, accouchement dans l’eau, absence de membres postérieurs fonctionnels, système reproducteur) ne peuvent pas être expliquées par des mutations graduelles. L’oreille interne des cétacés, par exemple, est spécialisée pour l’audition sous-marine et n’a aucun équivalent progressif connu chez les prétendus ancêtres terrestres. Même les soi-disant intermédiaires comme Ambulocetus ne montrent pas de véritables traits transitionnels, mais plutôt des combinaisons de traits mosaïques1.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
Les changements observés sont des adaptations dans des créatures déjà créées avec un potentiel de variation, mais il n’y a aucune preuve que ces animaux se sont transformés les uns vers les autres. L’idée que des animaux semi-aquatiques évoluent en baleines n’est jamais observée, mais seulement inférée.
D’après le scénario évolutionniste, la transition aurait eu lieu en quelques millions d’années seulement. Les milliers de mutations bénéfiques nécessaires, affectant de nombreux systèmes à la fois (locomotion, respiration, reproduction…), n’ont mathématiquement pas le temps de s’accumuler. De plus les mutations, qui sont des erreurs de réplication du code, sont neutres ou nuisibles à l’organisme. On les imagine difficilement transformer une créature terrestre en aquatique. Au contraire les mutations abîment continuellement le génome des organismes.
Les illustrations populaires (dans les manuels ou musées) présentent une progression linéaire et convaincante, mais ces reconstitutions sont hautement spéculatives et parfois contredites par de nouvelles découvertes (ex. Rodhocetus, supposé avoir des nageoires caudales mais dont on a découvert ensuite des os du bassin compatibles avec la marche terrestre).
Il n’existe aucune preuve génétique démontrant un tel changement progressif. Toute affirmation repose sur une interprétation phylogénétique des ressemblances et non sur une démonstration empirique de transformation d’un génome terrestre en un génome de baleine.
Pakicetus (« la baleine du Pakistan »)
Pakicetus était un animal terrestre ressemblant à un loup ou un chien. Il est loin de ressembler à une une baleine en devenir. L’argument principal en faveur d’un lien avec les baleines repose sur une caractéristique de l’oreille moyenne (bulla tympanique) partagée avec les cétacés. Mais ce trait est insuffisant pour prouver un lien évolutif, il peut simplement refléter une ressemblance fonctionnelle ou de design. Partager un os ne ferait pas de Pakicetus une baleine.
L’anatomie complète de Pakicetus montre qu’il marchait sur la terre ferme, ne présentait aucun signe de vie aquatique avancée (pas de nageoires, queue de baleine, souffle, etc.) et aurait bu de l’eau douce, selon l’analyse isotopique des dents. Il vivait près de l’eau, comme un cerf ou un crocodile aujourd’hui, cela ne le transforme pas en ancêtre des baleines.
Les manuels et médias ont largement diffusé des images incorrectes de Pakicetus comme une semi-baleine amphibie. Ces représentations ont été maintenues même après les découvertes prouvant qu’il était terrestre. Les chercheurs ont fini par conclure dans Nature:
« Pris dans leur ensemble, les caractères du crâne indiquent que les pakicétidés étaient terrestres, et que le squelette locomoteur présente des adaptations à la course. … Les pakicétidés étaient des mammifères terrestres, pas plus amphibies qu’un tapir. »
Spoor, F., Bajpai, S., Hussani, S., Kumar, K., Thewissen, J., Vestibular evidence for the evolution of aquatic behavior in early cetaceans, Nature, 2001.
Sur l’oreille interne, au final:
« Le plus ancien d’entre eux, Ichthyolestes [faisant référence à Pakicetus], montre une taille de canal similaire à celle des plus proches parents vivants des cétacés, les artiodactyles. »
Les artiodactyles sont les mammifères ongulés à nombre pair de doigts:
- cochons, hippopotames, chameaux, lamas, cerfs, girafes, antilopes, moutons, chèvres, bovins …
- Ce groupe n’inclut pas les baleines (selon la classification traditionnelle).
Dans l’oreille interne, les canaux semi-circulaires aident à l’équilibre et au contrôle des mouvements. Leur taille et leur forme reflètent souvent le mode de vie locomoteur (ex. course terrestre, nage, etc.). Chez les cétacés modernes, ces canaux sont très réduits, adaptés à la nage fluide sous-marine, il y a moins de besoin de contrôle fin des mouvements rapides de la tête comme chez les coureurs terrestres.
L’étude de Nature (2001) a montré que les canaux semi-circulaires de Pakicetus étaient de taille similaire à ceux des artiodactyles terrestres (cochons, hippopotames, cerfs…). Ils ne sont pas réduits comme ceux des baleines! Pakicetus était probablement un coureur terrestre, pas un nageur aquatique.
Ambulocetus (« la baleine qui marche »)

Malgré l’image ci-dessus, Ambulocetus n’a jamais été retrouvé complet. Le squelette est partiel, avec plusieurs zones importantes manquantes ou reconstituées par inférence. Le fossile original (Thewissen et al., 1994) comprend:
- Une partie du crâne
- Des vertèbres (cervicales, thoraciques, lombaires)
- Des côtes
- Une patte avant partielle
- Une portion des membres postérieurs, mais sans les pieds (pattes/phalanges distales) complets
- Aucune trace de la queue (ni vertèbres caudales, ni nageoire)
Ambulocetus est souvent représenté avec des pieds palmés, une queue partiellement transformée en nageoire (comme chez une loutre ou une baleine archaïque) et un mode de nage en « ondulation verticale ». Mais les pieds ne sont pas connus, donc l’existence de membranes palmées est une supposition artistique, non une observation fossile. Même l’auteur de la découverte, Hans Thewissen, admet que certaines caractéristiques ont été extrapolées.2
Ambulocetus est présenté comme une créature semi-aquatique intermédiaire, un amphibien entre les mammifères terrestres et les baleines, capable de marcher et de nager.
Le squelette a été reconstitué à partir de fossiles partiels, notamment sans le bassin, ce qui rend incertaine l’évaluation de sa locomotion. Il est spéculatif d’affirmer qu’il nageait comme une baleine alors qu’on ne connaît pas précisément la forme de la colonne, du bassin, ou de la queue.
Les premières illustrations montraient Ambulocetus avec une queue de baleine, alors que rien dans les fossiles ne justifie cela. Un animal avec une queue de crocodile et des pattes de loutre a été imaginé sans preuve suffisante.
La structure de la mâchoire, des dents, des vertèbres suggère un animal terrestre avec certaines adaptations à la nage et pas un cétacé. Il n’a pas de caractéristiques propres aux cétacés modernes: pas de souffle, pas d’oreille interne spécialisée pour l’audition sous-marine. Ce n’est pas une baleine en transition, c’est un mammifère terrestre qui pouvait nager.
Rodhocetus (« Baleine de Rodh »)

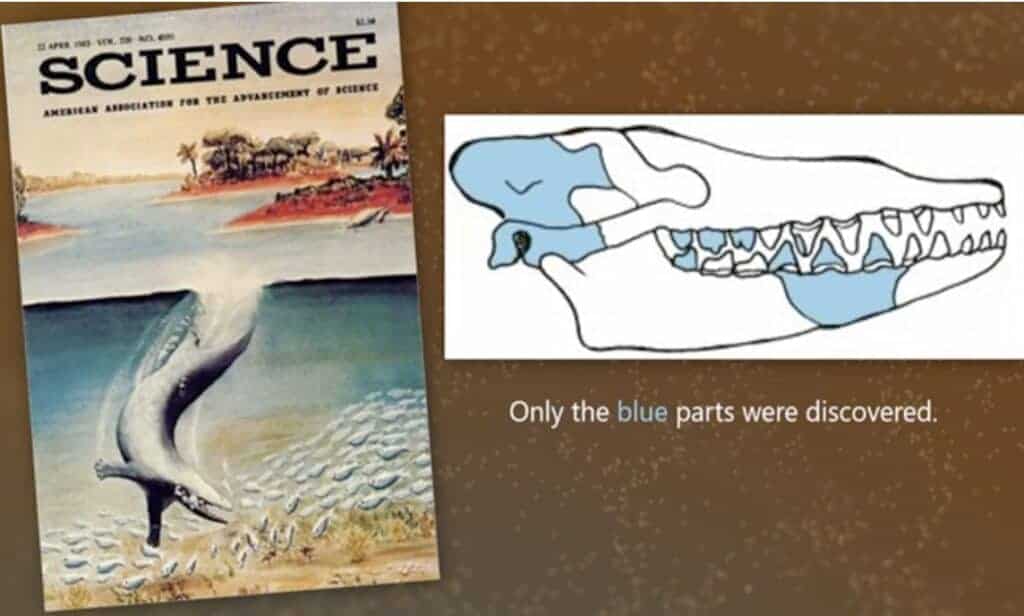
Voici une autre « Baleine du Pakistan » (en référence à la région de la Rodh Valley au Pakistan). L’image initiale était sublime mais complètement fantaisiste.34 Le squelette retrouvé en 1994 (Gingerich et al.) comprenait:
- Une partie du crâne
- Des côtes
- Des vertèbres thoraciques et lombaires
- Des os du bassin (ischium, ilium) et une portion de l’articulation avec les pattes postérieures
- Aucune trace de la queue ni des pattes ni des nageoires avant!
Une déclarations de Philip Gingerich (paléontologue évolutionniste dans PBS Nova a beaucoup retenu l’attention:
« We don’t have the fluke. We don’t have the flippers. »
« Nous n’avons pas la nageoire caudale. Nous n’avons pas les nageoires (avant). »
« Si vous n’avez pas de nageoires antérieures, je ne pense pas que vous puissiez avoir une nageoire caudale en forme de fluke [comme chez les baleines] et une nage réellement propulsée, et j’ai donc désormais des doutes sur le fait que Rodhocetus ait eu une nageoire caudale. »
C’est Gingerich lui-même (le découvreur) qui a corrigé les affirmations précédentes. Il avait imaginé une nageoire caudale (comme une baleine) sur la base de quelques vertèbres et avait supposé des nageoires avant, sans os fossiles pour les confirmer. Il a reconnu plus tard que le bassin de Rhodocetus était solide et articulé à la colonne, ce qui implique qu’il pouvait marcher. Le dessin d’origine (difficile à retrouver) a donc été complètement revu.
Gingerich ne pense pas que Rodhocetus a une queue puissante et une nage propulsée. Le squelette s’arrête au sacrum (la région pelvienne). Cela veut dire qu’on ne connaît rien de la queue de Rodhocetus (ni sa longueur, ni sa puissance, ni son extrémité).
Le Dr Jerry Bergman écrit dans son étude critique sur l’évolution de la baleine:
« Il n’existe aucune preuve d’une quelconque structure de nageoire caudale (fluke) dans les fossiles connus de Rodhocetus. Il lui manque même les vertèbres en forme de boule nécessaires pour soutenir un fluke — ce qui pose un problème majeur puisque tous les cétacés possèdent ces vertèbres sphériques et une nageoire caudale. »
Whale Evolution: A Whale of a Tale – Creation Research Society Quarterly 2012. 49:122–134.
Dans les premières reconstructions affichées dans les musées, on avait représenté Rodhocetus avec une queue de type baleine, des nageoires palmées et un évent en formation. Or Gingerich lui-même a reconnu plus tard que ces éléments étaient supposés et non basés sur les fossiles retrouvés. Sur la base de quelques fragments de Rodhocetus, on peut très bien le voir comme un mammifère amphibie (type loutre ou phoque ou Desman de Moscovie) mais pas une “proto-baleine”.
Vidéo courte sur le Desman de Moscovie (où on voit notamment son type de nage, bien différencié de celui d’une baleine):
https://www.youtube.com/watch?v=N3BAWMndqEo&ab_channel=YuriIvlev
Sur la page « évolutionniste » de wikipédia de Rodhocetus on peut lire:
« Par une analyse en composantes principales, Gingerich (2003) a démontré que Rodhocetus avait des proportions du tronc et des membres similaires à celles du desman de Russie, un nageur propulsé par ses pattes utilisant principalement sa queue comme gouvernail. À partir de cela, Gingerich a conclu que Rodhocetus nageait surtout en surface par des battements alternés de ses pattes postérieures, et qu’il était isolé par de la fourrure plutôt que par du lard, comme c’est le cas chez Dorudon et les cétacés modernes, ce qui le rendait flottant et incapable de plonger en profondeur. »

Desman et Rodhocetus partagent un profil morphologique qui suggère une nage par propulsion des pattes arrière, avec une queue servant de gouvernail, et non une nage type “baleine” avec fluke. Quand on dit que la queue sert de gouvernail, cela signifie que la queue ne fournit pas la propulsion principale (elle n’est pas une nageoire caudale puissante comme chez les dauphins ou thons), mais elle sert à orienter, stabiliser et diriger l’animal pendant la nage, un peu comme une “barre de direction” qui permet de virer à gauche/droite, de maintenir la stabilité, ou d’ajuster la trajectoire.
Chez la baleine, la propulsion principale est contrôlée par les muscles puissants du tronc et de la queue qui déplacent le fluke de haut en bas (et non latéralement comme chez les poissons). C’est le moteur principal de la nage. La direction, ce n’est pas le fluke qui dirige, mais les nageoires pectorales (flippers), qui agissent comme des gouvernails ou des ailes d’avion. La petite nageoire dorsale et la forme du corps aident aussi à la stabilité.
En fait, baleine et rodhocetus sont complètement opposés dans leur fonctionnement natatoire.
- La queue sert à diriger chez rodhocetus, chez la baleine elle sert de moteur, de propulsion.
- Les membres antérieurs, chez rodhocetus, servent à marcher et secondairement à se propulser dans l’eau (voir vidéo sur le Desman), chez la baleine ils ont le rôle de gouvernail.
- Rodhocetus nage en surface, et ne peut pas aller en profondeur, la baleine plonge à des centaines de mètres de profondeur
| Fonction | Rodhocetus (selon Gingerich 2003) | Baleine moderne |
|---|---|---|
| Propulsion principale | Pattes postérieures (battements alternés) | Fluke (queue horizontale puissante) |
| Rôle de la queue | Gouvernail (stabiliser, orienter) | Moteur (propulsion principale) |
| Rôle des membres antérieurs | Probablement marche/nage secondaire | Flippers = gouvernails et stabilisateurs |
| Isolation thermique | Fourrure (comme loutres/desmans) | Lard (blubber) |
| Mode de vie | Semi-aquatique, surface swimmer, incapable de plonger profond | Entièrement aquatique, plongeur capable de centaines de mètres |
J’alerte également sur le problème reproductif dans l’idée de l’évolution d’un mammifère ayant les testicules externes (en dehors du corps) par rapport aux baleines où les organes reproducteurs sont internes (près des reins, bien à l’intérieur du corps).
Les spermatozoïdes exigent une température plus basse que la température corporelle.La solution chez les mammifères terrestres (incluant probablement Rodhocetus) est la descente des testicules hors du corps pour maintenir la spermatogenèse. Si on les migre à l’intérieur, ils seraient trop chauds et stériles.
Chez les baleines, la solution est un système de refroidissement sophistiqué (contre-courant sanguin entre artères et veines, parfois relié aux nageoires pectorales qui servent de “radiateurs”).
Le passage du système reproducteur externe à interne exige la relocalisation anatomique complète des organes reproducteurs. Il faudrait une évolution simultanée d’un système vasculaire de refroidissement hyper spécialisé, sans quoi la reproduction serait impossible. Une transition “incomplète” serait non viabl, un mâle avec testicules internes mais sans système de refroidissement serait stérile.
Le problème reproductif est central, la transformation “testicules externes/internes avec refroidissement” n’a aucune documentation fossile et représente une rupture anatomique et physiologique majeure.
Rodhocetus n’était pas une “baleine” mais un mammifère semi-aquatique de taille moyenne, plus proche par la taille et l’écologie d’un phoque ou d’une loutre géante. Phoques, loutres et desmans ont les organes reproducteurs externes (dans un scrotum). Seules les baleines et dauphins présentent le système interne avec refroidissement, ce qui pose un problème évolutif majeur: il n’existe pas de formes fossiles documentant cette transition.

Pour raconter une histoire de transition convaincante, les artistes avaient représenté Rhodocetus comme une créature déjà très « baleinesque », avec une queue de cétacé et des membres avant transformés en nageoires. Ces représentations ont persisté dans les manuels et documentaires, même après les aveux de Gingerich.
Le Dr Chris Rupe écrit au sujet de Rodhocetus:
« Ce qui est intéressant, c’est que les restes fossiles de la découverte originale ne présentent aucune trace fossile ni des membres antérieurs et postérieurs, ni des os de la queue — y compris la vertèbre sphérique, principal indicateur d’un fluke présent chez toutes les baleines modernes.
Il s’avère que tous les caractères squelettiques distinctifs essentiels manquent. Il n’y a aucune preuve d’un grand ensemble queue/fluke, aucune preuve d’une queue massive servant à la propulsion, aucune preuve de membres postérieurs réduits, aucune preuve de fusion de la patte arrière dans le ventre, aucune preuve de nageoires antérieures, aucune preuve d’une musculature cervicale massive et profilée, et aucune preuve qu’une position de nage était sa posture naturelle. »
Tales of Walking Whales – page 8,9
Rodhocetus a été surinterprété pour appuyer le scénario évolutif ; il s’agit probablement d’un mammifère amphibie bien conçu et non d’un cétacé. L’image initiale était très éloignée de ce qu’il était réellement mais le public a été laissé dans l’impression que la progression évolutive vers la baleine était frappante.
Le fait qu’il ait un bassin solide et articulé à la colonne vertébrale indique une locomotion terrestre, ce qui est contraire à ce qu’on attend d’une « quasi-baleine ». Son utilisation comme preuve forte d’évolution est un exemple de science narrative basée sur des attentes plutôt que sur des données objectives.
Basilosaurus (« roi des lézards »)

Basilosaurus serait un « proto-cétacé » très aquatique, avec une forme serpentine et de petits membres postérieurs atrophiés. Même les évolutionnistes reconnaissent aujourd’hui que Basilosaurus est une branche latérale, non un ancêtre direct. Il est trop spécialisé, avec une forme serpentine et des comportements uniques. Thewissen et al. (2009) reconnaissent que Basilosaurus n’est pas dans la lignée directe des baleines modernes.
Il est souvent présenté comme preuve d’un ancêtre terrestre, mais ces membres sont trop petits pour la locomotion, ils étaient probablement utilisés pour l’accouplement (comme l’admettent certains paléontologues).
Chez les baleines, le trou nasal (spiracle) est situé tout en haut du crâne. Chez Basilosaurus, la narine reste positionnée à l’avant, ce qui pose problème pour une respiration aquatique optimisée. Il n’y a pas l’étape clé de la migration du nez vers le sommet du crâne.
Si l’on met Ambulocetus, Rodhocetus et Basilosaurus en série, on n’observe pas une gradation linéaire progressive, mais des formes distinctes.
Les manuels montrent souvent Basilosaurus avec:
- Un corps lisse et profilé comme une baleine moderne,
- Une nageoire caudale horizontale (fluke),
- Des nageoires avant bien développées.
Le problème est qu’il n’y a aucune preuve fossile directe d’une nageoire caudale. La forme serpentine et ondulante rappelle plus un serpent marin géant qu’une baleine. Basilosaurus nageait probablement en ondulant tout son corps, contrairement aux cétacés modernes.
L’anatomie du Basilosaurus n’indique pas une transition mais une forme aboutie. Il ne s’agit pas d’une créature en cours de transformation, mais d’un animal aquatique accompli. Les éléments clés censés prouver une origine terrestre sont soit absents, soit mal interprétés. L’idée qu’il représente une étape dans l’évolution des baleines repose davantage sur des inférences que sur des faits démontrés.
Dorudon
Dorudon est un cétacé fossile de la famille des basilosauridés, tout comme Basilosaurus. Il vivait à la fin de l’Éocène (environ 40 à 34 millions d’années selon la chronologie évolutionniste) et était entièrement aquatique, avec des membres postérieurs atrophiés.
Dorudon, tout comme Basilosaurus a un corps long et serpentin (ondulant), il possède de petites pattes postérieures, trop petites pour permettre la marche sur terre et est donc totalement adapté au milieu marin. Ce n’est pas une forme de « transition » en train de s’adapter à l’eau. C’est déjà un animal marin abouti.
Même des scientifiques évolutionnistes ont reconnu que les basilosauridés (dont Dorudon) ne sont pas des ancêtres directs des baleines modernes parce qu’il a une forme serpentine trop distincte, des dents (« cheek teeth ») très particulières, sans lien clair avec les mysticètes (baleines à fanons) ou les odontocètes (baleines à dents). Selon la paléontologue Barbara J. Stahl:
« La forme serpentine du corps et la forme particulière des dents jugales montrent clairement que ces archéocètes [les basilosauridés] ne pouvaient absolument pas être les ancêtres des baleines modernes. »
Stahl, B., Vertebrate History: Problems in evolution, McGraw-Hill, p. 489, 1974,
Les membres postérieurs ne servent pas à marcher, ils sont trop petits pour la locomotion et étaient probablement utilisés pour l’accouplement (comme « bras d’accouplement »). Donc ces membres ne sont pas des vestiges de jambes en train de disparaître, mais des structures fonctionnelles spécifiques, comme chez certains reptiles ou poissons.
Certains paléontologues pensent que Dorudon vivait en même temps que des baleines à fanons primitives. Donc il ne peut pas être leur ancêtre direct, puisqu’ils auraient été contemporains. Dorudon est en fait un exemple d’animal marin pleinement formé et non une créature en transition depuis la terre ferme vers la mer.
Conundrum avec la génétique
Les manuels et les documentaires évolutionnistes présentent généralement la chose comme ceci:
Pakicetus -> Ambulocetus -> Rodhocetus -> Basilosaurus -> Baleines modernes
Ce récit est basé sur la morphologie (os, dents, crânes, membres, etc.), dans une vision linéaire d’évolution progressive.
Dans les années 1990–2000, des études moléculaires (ADN, protéines) ont montré que les cétacés sont proches génétiquement des hippopotames. Ils sont inclus dans les Artiodactyla (ongulés à doigts pairs : cerfs, cochons, etc.). Aujourd’hui, les évolutionnistes disent que les baleines sont un sous-groupe des artiodactyles, et leur plus proche cousin vivant est… l’hippopotame.
Ils disent également que Pakicetus, Ambulocetus et Rodhocetus sont des cétacés primitifs mais ces cétacés primitifs font partie des artiodactyles. Donc l’hippopotame et les baleines auraient un ancêtre commun artiodactyle terrestre et Pakicetus et Ambulocetus font partie de la branche cétacéforme de cet ancêtre.
Il y a toutefois un problème:
- Morphologiquement, Pakicetus ressemble à un mesonychia (un groupe entièrement éteint de mammifères carnivores à sabots) et pas à un artiodactyle.
- L’hippopotame ne ressemble en rien à Pakicetus ou à une “baleine en devenir”.
Historiquement, les évolutionnistes pensaient que les baleines descendaient des mesonychia, car leurs crânes et dents sont similaires à ceux des cétacés primitifs (comme Pakicetus). Ernst Mayr a déclaré en 2001:
« Il existe également une magnifique série de stades intermédiaires entre les ongulés mésonychidés et leurs descendants, les baleines. »5
Kenneth Rose et d’autres expliquent maintenant que les similitudes entre les baleines et les mésonychidés se sont produites indépendamment dans les deux groupes en raison d’une « évolution convergente ».
Le Dr Randy Guliuzza commente:
« La convergence n’est pas une observation découlant de causes objectivement discernables. C’est en réalité une affirmation fondée sur des représentations mentales d’organismes divers évoluant vers des traits similaires, façonnés au fil du temps par des pressions environnementales similaires — lesquelles ne sont pas de véritables pressions quantifiables, mais n’existent qu’en tant que figures de style. »6
La génétique dit que les baleines sont proches des hippopotames mais la morphologie crânienne et dentaire dit que les baleines ressemblent plus aux mesonychia (pas aux artiodactyles classiques). Cela crée une tension:
- Soit on dit que les ressemblances crâniennes sont dues à l’évolution convergente (donc non issues d’un ancêtre commun)
- Soit on dit que les artiodactyles ont perdu ces traits, ce qui implique des transformations supplémentaires ad hoc
Une citation évolutionniste présente ce problème:
« Malgré cette preuve que les cétacés [baleines] ont évolué à partir des artiodactyles [mammifères à doigts pairs comme les cerfs, les moutons et les cochons], d’importantes divergences subsistent. Si les cétacés appartiennent aux artiodactyles, alors les similarités dans la morphologie crânienne et dentaire entre les mesonychia [mammifères carnivores éteints] et les cétacés doivent être le résultat d’une évolution convergente, ou bien ces traits ont dû être perdus chez les artiodactyles. En outre, les données moléculaires favorisent une relation de groupe frère entre les baleines et les hippopotames. Cela entre en conflit avec la vision conventionnelle fondée sur la morphologie, selon laquelle les hippopotames sont plus proches des autres artiodactyles que des baleines. »
Rose, K. D. 2001. Evolution: The Ancestry of Whales. Science. 293 (5538): 2216-2217.
Selon le modèle créationniste, les similitudes entre les baleines, les mesonychia et les hippopotames ne s’expliquent pas par une ascendance commune, mais par un Concepteur en commun ayant appliqué des principes fonctionnels partagés selon les besoins des créatures. Des traits morphologiques ou génétiques similaires peuvent refléter des fonctions analogues (comme la vie aquatique) sans impliquer une parenté.
Le Dr Chris Rupe écrit:
« Bien que les théoriciens de l’évolution des baleines se sentent obligés d’argumenter que les baleines proviennent d’animaux à sabots, cela est très problématique car les sabots ne sont pas utiles pour nager et ne peuvent pas se transformer en pieds palmés avec des os en forme de doigts.
Les évolutionnistes pensent que les animaux à sabots ont évolué en sabots fusionnés pour remplacer les cinq orteils communs à la plupart des mammifères. C’est un problème car ni Pakicetus, ni les deux autres formes [Rodhocetus et Ambulocetus] transitionnelles réputées n’ont de sabots.
Pakicetus montre qu’il avait des orteils, pas des sabots, tout comme Ambulocetus (les extrémités de Rodhocetus manquaient complètement et ne représentent qu’une invention artistique).
Indohyus, le plus récemment découvert “cétacé primitif” semblable à un petit chevreuil, est censé avoir immédiatement précédé Pakicetus, et il lui manque aussi des sabots évidents — mais l’artiste a ajouté de petits sabots au bout des orteils (voir l’image de Nature 2007 ci-dessous, figure 11)…
D’un côté, les similitudes génétiques suggèrent que les baleines auraient évolué d’une lignée ancestrale à sabots, et pourtant les preuves fossiles des prétendus précurseurs des baleines ne correspondent pas à cette hypothèse. »
Contrairement à l’évolution qui regroupe des espèces sur la base de ressemblances partielles, la classification créationniste s’appuie sur un continuum reproductif, morphologique et fonctionnel, selon lequel baleines, hippopotames et mesonychia appartiennent à des baramins distincts, sans liens généalogiques entre eux.
Pour les créationnistes :
- Les cétacés forment un baramin distinct (non lié aux hippopotames ni aux mésonychidés),
- Les mesonychia sont probablement un autre baramin terrestre carnivore éteint,
- Les hippopotames appartiennent au baramin Hippopotamidae.
Darwin et l’évolution de la baleine
A citer les darwinistes modernes, on en oublierait presque la théorie initiale de Darwin lui-même sur les baleines:
« …nageant pendant des heures, la gueule largement ouverte, attrapant ainsi, comme une baleine, des insectes dans l’eau. Même dans un cas aussi extrême que celui-ci, si l’approvisionnement en insectes était constant et si des concurrents mieux adaptés n’existaient pas déjà dans le pays, je ne vois aucune difficulté à ce qu’une race d’ours soit rendue, par sélection naturelle, de plus en plus aquatique dans sa structure et ses habitudes, avec des gueules de plus en plus grandes, jusqu’à ce qu’une créature aussi monstrueuse qu’une baleine soit produite. »
Darwin, C. 1859. The Origin of Species by Means of Natural Selection. London: John Murray, 184.
Conclusion
Ces fossiles de Pakicetus, Ambulocetus, Rodhocetus, Basilosaurus et autres, ne démontrent pas un processus d’évolution vers les baleines. Ils sont des créatures uniques et adaptées, interprétées a posteriori selon un paradigme évolutionniste et reconstituées avec beaucoup d’imagination et peu de preuves directes. Le récit évolutionniste est forcé pour produire un scénario « gradualiste ».

Le souffle de la baleine est un exemple frappant de conception. Contrairement aux autres mammifères, elle respire de l’air tout en vivant dans l’eau. Son évent est placé exactement au sommet de la tête : un emplacement parfait qui lui permet de respirer rapidement sans sortir tout son corps.
Cet évent possède aussi un clapet musculaire qui le rend étanche pendant la plongée et s’ouvre instantanément quand la baleine refait surface. Un système d’une précision telle que la baleine ne pourrait survivre si un seul élément manquait. Ces évents au sommet du crâne sont complètement absents chez Pakicetus, Ambulocetus et Rodhocetus.
Les mutations aléatoires ne peuvent pas expliquer un mécanisme aussi complexe et précis. Vu à travers la création, le dessein est évident. Dieu a créé les baleines entièrement adaptées à la vie marine dès le commencement.
C’est la même pour le système d’écholocation. Les candidats Pakicetus, Ambulocetus, Rodhocetus et autres n’en montrent aucun signe (pas de melon, pas de lèvres phoniques, pas de système auditif découplé, ni de mandibule creuse spécialisée.). C’est pourtant l’une des caractéristiques les plus complexes et impressionnantes des cétacés (notamment les odontocètes, comme les dauphins et les cachalots).
L’écholocation chez les cétacés modernes repose sur un système intégré extrêmement sophistiqué:
| Composant | Fonction |
|---|---|
| Melon (tissu adipeux frontal) | Focalise les ondes sonores avant l’émission |
| Phonic lips (lèvres phoniques) | Génèrent les clics ultrasoniques |
| Os tympaniques découplés | Isolation acoustique de chaque oreille |
| Mandibule creuse + coussinet adipeux | Conduit les sons vers l’oreille interne |
| Cerveau (traitement rapide du signal) | Analyse des échos pour modéliser l’environnement |
Ce système n’a d’équivalent chez aucun autre mammifère terrestre (le système des chauves-souris est profondément différent).
L’écholocation est un système irréductiblement complexe:
- Aucun fossile connu ne montre l’émergence progressive de ce système.
- Tous les composants doivent être présents et fonctionnels simultanément, sinon l’écholocation ne marche pas.
- Il n’existe aucun exemple intermédiaire fonctionnel dans les archives fossiles.
La « baleine du Pakistan », la « baleine qui marche » et la baleine de la vallée de Rodh » n’étaient en fait aucunement « baleinesque ». Ils étaient des organismes « terrestres » avec pour certains peut être quelques adaptations à la nage, comme c’est le cas pour de nombreux organismes aujourd’hui qui peuvent explorer deux terrains. Les trois ont trop rapidement été qualifiés de « baleines » mais les données ultérieures ont contredit cette idée.
Basilosaurus et Dorudon étaient eux des « serpents de mer » et fonctionnaient complètement différemment des baleines.
Les évolutionnistes disent maintenant que les baleines et les hippopotames partagent un ancêtre commun dans le cadre de l’ordre des artiodactyles (ongulés à doigts pairs). Mais cet ancêtre commun n’a jamais été observé. Il est hypothétique. Ce lien est basé uniquement sur des similarités génétiques et non sur des fossiles montrant une transition progressive.
Il n’existe aucune série fossile continue allant d’un artiodactyle ressemblant à un hippopotame vers une forme progressivement aquatique. Les fossiles utilisés pour illustrer cette transition (Pakicetus, Ambulocetus, Rodhocetus) ressemblent davantage à des mammifères carnivores terrestres (comme les mésonychidés) qu’à des artiodactyles.
Pour faire correspondre la génétique et la paléontologie, les évolutionnistes ont décidé de reclasser rétroactivement Pakicetus et les autres:
- Ils disent maintenant que Pakicetus n’est plus un mésonychidé, mais un artiodactyle basal (même s’il n’en a pas les caractéristiques).
- Les similarités avec les mésonychidés sont attribuées à une évolution convergente (ressemblances sans lien généalogique ce qui est plutôt cohérent avec le design modulaire utilisé à plusieurs reprises par un concepteur en commun).
Cela revient à sauver le modèle en complexifiant le récit, sans preuves transitionnelles cohérentes.
Certains évolutionnistes reconnaissent la nature ténue de ce scénario évolutif:
« [Les baleines à fanons] sont apparues durant l’Éocène supérieur, mais les preuves de leur évolution précoce demeurent à la fois rares et controversées. »7
Dans le modèle créationniste, les baleines ont toujours été des baleines, les oiseaux des oiseaux, les poissons des poissons. Il y a potentiellement eu de la variation au sein des genres, mais pas de transition d’un genre à l’autre.
La génétique, loin de confirmer l’évolution progressive des baleines, a :
- Contredit la version morphologique traditionnelle
- Créé un conflit interne entre données fossiles et moléculaires
- Forcé des réinterprétations ad hoc pour préserver le récit évolutif
Du point de vue créationniste, cette confusion illustre qu’il ne s’agit pas d’un modèle vérifiable, mais d’un narratif ajustable, fondé sur des hypothèses évolutives plutôt que sur des continuités reproductives, morphologiques et fonctionnelles réelles.
Le site Real Science Radio résume et fournit une liste des problèmes du scénario de l’évolution de la baleine:
Dans l’article suivant, j’explore le défi insurmontable à relever concernant l’évolution de la baleine à partir d’un mammifère terrestre:
Références:
- Traits mosaïques = combinaison de caractéristiques de plusieurs groupes. Certains organismes vivent dans des environnements de transition (eau/terre, air/sol, chaud/froid, etc.) et possèdent donc une combinaison de traits adaptés à plusieurs milieux. C’est ce qu’on peut appeler une adaptation multifonctionnelle, éclectique. Cela ne prouve pas que l’un évolue vers l’autre.
- *Thewissen et al., Science (1994), « Skeletons of Ambulocetus natans ».
- https://creation.com/rodhocetus.
- Tales of Walking Whales – page 2 – Dr Chris Rupe liste des images de manuels scolaires présentant Rodhocetus avec nageoire caudales et nageoires antérieures.
- Mayr, E. 2001. What Evolution Is. New York: Basic Books, 63.
- Major Evolutionary Blunders: Are Whales and Evolution Joined at the Hip?
- – Fordyce, R. & F. Marx. 2018. Gigantism Precedes Filter Feeding in Baleen Whale Evolution. Current Biology.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!