

Phylums précoces, innovation éteinte: une lecture critique de la macroévolution
Un point très important, souvent passé sous silence dans les manuels d’évolution, mais mis en avant par plusieurs biologistes créationnistes et même certains paléontologues sceptiques du néo-darwinisme classique, est l’apparition des grands groupes (phyla) suivi des subdivisions à mesure que l’on remonte la colonne géologique. L’évolution, théoriquement, postule l’inverse, ce qui pose de sérieux problèmes.
L’apparition brutale des grands groupes dans le registre fossile
| Niveau taxonomique | Apparition dans le registre fossile |
|---|---|
| Embranchements (phyla) | Très tôt, souvent dès le Cambrien |
| Classes | Peu après, encore dans le Cambrien ou Ordovicien |
| Ordres | Progressivement après les classes |
| Familles, genres, espèces | Tardifs, en grande partie après les groupes supérieurs |
Dans l’explosion cambrienne, on observe l’apparition brutale de la plupart des plans corporels majeurs, comme les arthropodes, mollusques, chordés, etc. Ces phylums sont entièrement formés, avec aucune forme ancestrale en dessous.
Dans le modèle évolutionniste traditionnel on s’attendrait à ce que l’évolution débute par des formes simples, puis que les divisions apparaissent progressivement par diversification:
- espèces → genres → familles → ordres → classes → phyla.
Mais le registre fossile montre l’inverse les phyla apparaissent en premier, puis les subdivisions se développent ensuite dans les phylums déjà existants.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
C’est exactement ce que plusieurs modèles créationnistes baraminologiques avancent:
- Dieu aurait créé des types originels (baramins) avec une grande variabilité génétique intégrée.
- Par la suite, ces types se seraient diversifiés (adaptation, spéciation), donnant naissance à des espèces spécifiques au fil du temps.
- Cela n’implique pas l’ajout de nouvelles structures complexes, mais une spécialisation de la variabilité génétique disponible.
C’est une forme de variation intelligente et ciblée, orientée vers la spécialisation, et non vers l’innovation fonctionnelle (qui serait de la macro évolution).
Modèle évolutionniste — Ascendance commune
Dans le modèle évolutionniste classique, l’histoire du vivant est représentée comme un grand arbre phylogénétique dans lequel chaque groupe taxonomique (ordre, classe, phylum, etc.) descend d’un ancêtre commun hypothétique. Par exemple:
- tous les carnivores de l’ordre « Carnivora » descendraient d’un ancêtre commun carnivore primitif
- tous les mammifères de la classe « Mammalia » descendraient d’un ancêtre mammalien
- tous les vertébrés descendraient d’un poisson primitif
- en bout de chaîne il y aurait un ancêtre universel commun (LUCA)
Cette vision n’est imposée que par le modèle évolutionniste et non par l’observation. Il n’existe aucun fossile connu représentant ces « ancêtres communs » avec des traits intermédiaires pleinement convaincants pour chaque grande transition (ex. mammifère-reptile, reptile-oiseau).
Le modèle repose sur une extrapolation de similitudes morphologiques ou génétiques interprétées comme des preuves d’ascendance commune et suppose un mécanisme suffisant (mutations + sélection naturelle) pour produire des organes complexes entièrement nouveaux, ce que de nombreux critiques jugent non démontré expérimentalement.
GRANDS PHYLUMS ANIMAUX (≈ 35 phyla connus, en voici les principaux)
| Phylum | Description | Exemples |
|---|---|---|
| Chordata | Animaux avec une corde dorsale (au moins à un stade) | Vertébrés (poissons, amphibiens, reptiles, oiseaux, mammifères) |
| Arthropoda | Exosquelette, pattes articulées, corps segmenté | Insectes, araignées, crustacés |
| Mollusca | Corps mou, souvent avec coquille | Escargots, pieuvres, moules |
| Annelida | Vers segmentés | Lombrics, sangsues |
| Platyhelminthes | Vers plats | Ténias, douves |
| Nematoda | Vers ronds | Ascaris, oxyures |
| Echinodermata | Symétrie radiale chez les adultes, endosquelette | Étoiles de mer, oursins |
| Cnidaria | Corps gélatineux avec cellules urticantes | Méduses, coraux, anémones |
| Porifera | Pas de vrais tissus, filtreurs | Éponges |
| Bryozoa | Colonies aquatiques fixées | Bryozoaires |
| Ctenophora | Peignes ciliés, ressemblant aux méduses | Cténophores (peignes de mer) |
Dans le référentiel évolutif, tous ces grands phylums apparaissent entre 600 et 480 millions d’années. Les phylums sont souvent vus comme des plans corporels fondamentaux, la difficulté pour les évolutionnistes est qu’ils apparaissent de manière soudaine dans le registre fossile (on appelle cela l’explosion cambrienne).
Le problème de l’explosion cambrienne
Les évolutionnistes mettent souvent en avant les milliards et centaines de millions d’années pour justifier l’évolution mais quand on y regarde de plus près, les évolutions doivent se produire en quelques millions d’années seulement. On le voit notamment avec les phylums d’animaux actuels: arthropodes, chordés, mollusques, échinodermes, annélides, etc. Ces groupes émergent sans ancêtres fossiles clairs, et souvent avec une complexité morphologique avancée dès le départ. Il n’y a pas de formes transitionnelles incontestables observées.
Selon Darwin et ses successeurs, l’évolution devait produire de petits changements progressifs sur de longues périodes. Les groupes supérieurs (phyla) devaient émerger après des millions d’années de diversification graduelle: Espèces → genres → familles → ordres → classes → phylums. La plupart des plans corporels majeurs apparaissent à ce moment sans prédécesseurs fossiles évidents mais le registre fossile montre l’inverse:
- Les phylums apparaissent d’un coup, puis les classes, ordres, espèces apparaissent ensuite.
- La diversité morphologique maximale est au début, suivie d’une spécialisation descendante, non ascendante.
- La plupart des plans corporels majeurs apparaissent à ce moment sans prédécesseurs fossiles évidents.
Les évolutionnistes ont proposé plusieurs explications alternatives:
- Hypothèse de fossiles manquants, les ancêtres étaient mous, donc mal conservés.
- Gènes de développement (comme Hox), des petits changements génétiques auraient produit de grands effets.
- Hypothèse de l’évolution ponctuée (Gould & Eldredge), l’évolution se produit par sauts rapides, difficiles à capturer dans les fossiles.
- Des conditions géochimiques soudainement favorables à la fossilisation.
Ces idées sont souvent perçues comme ad hoc et n’expliquent pas pourquoi la complexité apparaît avant la diversité progressive attendue.
GRANDES CLASSES DU PHYLUM CHORDATA
| Classe | Groupe principal | Exemples |
|---|---|---|
| Mammalia | Mammifères | Humains, chats, baleines |
| Aves | Oiseaux | Moineaux, aigles |
| Reptilia | Reptiles | Serpents, crocodiles |
| Amphibia | Amphibiens | Grenouilles, salamandres |
| Actinopterygii | Poissons osseux à nageoires rayonnées | Truite, hareng |
| Chondrichthyes | Poissons cartilagineux | Requins, raies |
| Agnatha | Poissons sans mâchoires | Lamproies, myxines |
| Cephalochordata | Cordés non vertébrés | Amphioxus |
| Urochordata | Tuniciers | Ascidies |
Il est difficile de comprendre pourquoi des amphibiens et des reptiles sont restés des amphibiens et des reptiles sur des centaines de millions d’années mais qu’en même temps certains d’entre eux se sont transformés en des mammifères, des oiseaux etc…
On comprend mieux le registre fossile dans le cadre de la création suivie du déluge qui a emporté les organismes séquentiellement en fonction des terrains rencontrés (de la mer vers les terres). Basiquement 600 millions d’années du modèle évolutionniste s’expliquent dans l’année du déluge qui a créé les grandes caractéristiques géologiques catastrophiquement et fossilisé des trilliards d’organismes. Ce qu’il y a en dessous du Cambrien appartient globalement aux roches de la Création (excepté peut-être des spécificités comme la faune édiacarienne).
GRANDS ORDRES DE LA CLASSE MAMMALIA
| Ordre | Description | Exemples |
|---|---|---|
| Primates | Mains préhensiles, vision binoculaire | Humains, singes |
| Carnivora | Dents spécialisées, souvent carnivores | Loups, lions, ours |
| Cetacea | Aquatiques, pas de membres postérieurs | Baleines, dauphins |
| Rodentia | Incisives à croissance continue | Rats, écureuils |
| Chiroptera | Vol actif | Chauves-souris |
| Artiodactyla | Ongulés à nombre pair de doigts | Bœufs, moutons, cerfs |
| Perissodactyla | Ongulés à nombre impair de doigts | Chevaux, rhinocéros |
| Lagomorpha | Petits herbivores, deux paires d’incisives | Lapins |
| Sirenia | Aquatiques herbivores | Lamantins, dugongs |
| Proboscidea | Trompe, grandes oreilles | Éléphants |
| Monotremata | Pondent des œufs | Ornithorynque, échidné |
| Marsupialia | Poche abdominale | Kangourous, koalas |
Les classes ou ordres peuvent correspondre à des variations ou regroupements selon le design commun, mais ne traduisent pas une ascendance évolutive.
L’apparition soudaine des phylums dans les couches cambriennes est exactement ce qu’on attend d’une création par types distincts. Cela suggère que les grands plans corporels ont été créés séparément, puis ont diversifié ensuite par adaptation, spéciation, extinction. La discontinuité observée est un argument en faveur d’une origine non évolutive de la vie animale.
Rangs taxonomiques dans le modèle évolutionniste
Dans la classification biologique classique (système hiérarchique développé notamment par Carl von Linné), chaque organisme est classé selon un ensemble de rangs taxonomiques, du plus large au plus spécifique:
| Rang | Exemple (chien domestique – Canis lupus familiaris) |
|---|---|
| Domaine | Eukaryota (cellules avec noyau) |
| Règne | Animalia (animaux) |
| Embranchement / Phylum | Chordata (présence d’une corde dorsale) |
| Classe | Mammalia (mammifères) |
| Ordre | Carnivora (carnivores) |
| Famille | Canidae (chiens, loups, renards) |
| Genre | Canis |
| Espèce | Canis lupus |
| Sous-espèce | Canis lupus familiaris (chien domestique) |
Dans le postulat évolutionniste, chaque rang représente une branche dans l’arbre évolutif, issue d’un ancêtre commun avec les autres groupes de ce rang. Par exemple:
- Tous les mammifères (classe Mammalia) dériveraient d’un ancêtre mammalien commun.
- Tous les carnivores (ordre Carnivora) dériveraient d’un ancêtre carnivore commun, etc.
Perception créationniste des rangs taxonomiques
Le baramin est l’unité fondamentale de la création selon Genèse 1 : « selon leur espèce » (hébreu : min). Ce « type créé » ne correspond pas nécessairement à un rang taxonomique fixe. Il peut regrouper plusieurs espèces, voire genres.
| Rang | Accepté ? | Lecture créationniste |
|---|---|---|
| Domaine | oui | Purement descriptif (eucaryotes, procaryotes) |
| Règne | oui | Idem, mais non lié à une origine commune |
| Phylum | oui descriptif | Sert à classer selon les plans corporels, mais ne reflète pas une ascendance commune |
| Classe | oui partiellement | Des catégories utiles mais non forcément généalogiques |
| Ordre | oui | Peut contenir plusieurs types créés (ex. Carnivora = félins + canins) |
| Famille | possible baramin | Souvent équivalent au « type créé » (ex. Canidae, Felidae) |
| Genre | oui | Peut être trop spécifique pour représenter un type originel |
| Espèce | oui mais redéfini | Terme flou — l’espèce biblique ≠ espèce moderne (linéenne) |
Au-dessus du rang « famille » (ordre, classe, phylum), les preuves d’ancêtres communs deviennent hypothétiques ou fragmentaires. On ne retrouve pas de formes transitoires nettes entre, par exemple:
- Les félins et les canidés (ordre Carnivora),
- Les mammifères et les reptiles (classes),
- Les cordés et les arthropodes (phyla),
On parle alors de groupes-souches, de formes hypothétiques ou de mosaïques (ex. Tiktaalik, Archaeopteryx), mais jamais de séries continues vérifiables.
Pourquoi ces seuils « famille » et « genre » sont significatif?
Les hybridations fertiles se produisent souvent au sein de la famille ou du genre, et non entre différentes familles ou entre différents genres.
- Exemple avec le chien et le loup qui donnent une progéniture viable et fertile (même famille : Canidae),
- Le lion et le tigre donnent une progéniture viable mais partiellement fertile (même famille : Felidae),
- Le chien et le chat ne peuvent pas se reproduire, c’est impossible (familles différentes).
Les études baraminologiques montrent que la famille ou le genre (parfois un clade inférieur) sont souvent les frontières biologiques réelles.
Exemples concrets de baramins proposés
| Baramin (type créé) | Correspondance taxonomique approximative |
|---|---|
| Canin | Famille Canidae |
| Félin | Famille Felidae |
| Équin | Genre Equus (cheval, zèbre, âne) |
| Bovin | Tribu Bovini (bœuf, buffle, yack) |
| Anatidés (oiseaux d’eau) | Famille Anatidae |
| Palmipède marin (manchots, cormorans…) | Plusieurs ordres, un seul baramin possible selon certains |
| Humain | Homo sapiens, – type à part créé à l’image de Dieu (comprenant Erectus et Néandertal) |
| Groupe | Baramin proposé | Représentants modernes | Représentants fossiles possibles |
|---|---|---|---|
| Canidés | Baramin canin | Loup, chien, renard | Eucyon, Leptocyon (candidats, mais post-création) |
| Félidés | Baramin félin | Lion, tigre, chat | Panthera (divers fossiles) |
| Équidés | Baramin équin | Cheval, zèbre, âne | Hyracotherium |
| Proboscidiens | Baramin éléphantin | Éléphant d’Asie, d’Afrique | Mammouths, mastodontes |
| Ursidés | Baramin ursidé | Grizzly, ours polaire | Ursus spelaeus, etc. |
Conclusion
«Nous sommes toujours dans le brouillard par rapport à l’apparition des groupes majeurs d’organismes. Ils apparaissent dans le registre fossile comme Athéna qui sort de la tête de Zeus, intégralement et distinctement. C’est en contradiction avec la conception Darwinienne de l’évolution, à savoir le résultat d’une accumulation de millions de variations minuscules et graduelles.»
Jeffrey Schwartz, Sudden Origins: Fossils, Genes, and the Emergence of Species, p3
Dans la vision macroévolutionniste de nouveaux grands groupes devraient continuer à émerger, même lentement mais ce n’est pas ce qu’on observe, aucun nouveau phylum n’est apparu depuis le Cambrien. Cela s’accorde mal avec l’idée d’une innovation continue par mutations aléatoires.
Comme le disait Stephen Jay Gould, « L’explosion cambrienne produit non seulement de nouveaux groupes, mais la plupart des groupes majeurs existants » et Douglas Erwin (Smithsonian Institution) « La plupart des plans corporels animaux furent établis au Cambrien. Depuis, l’évolution s’est concentrée sur des variations internes. »
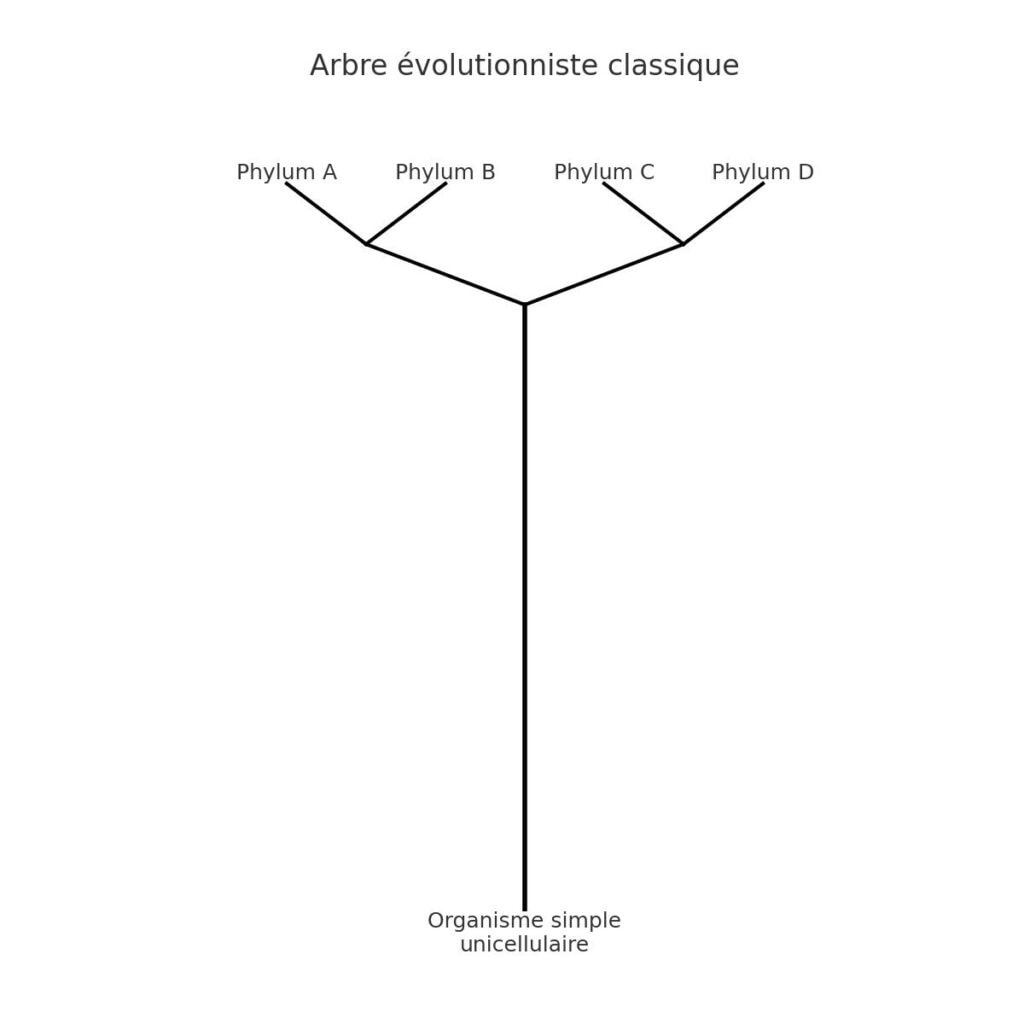
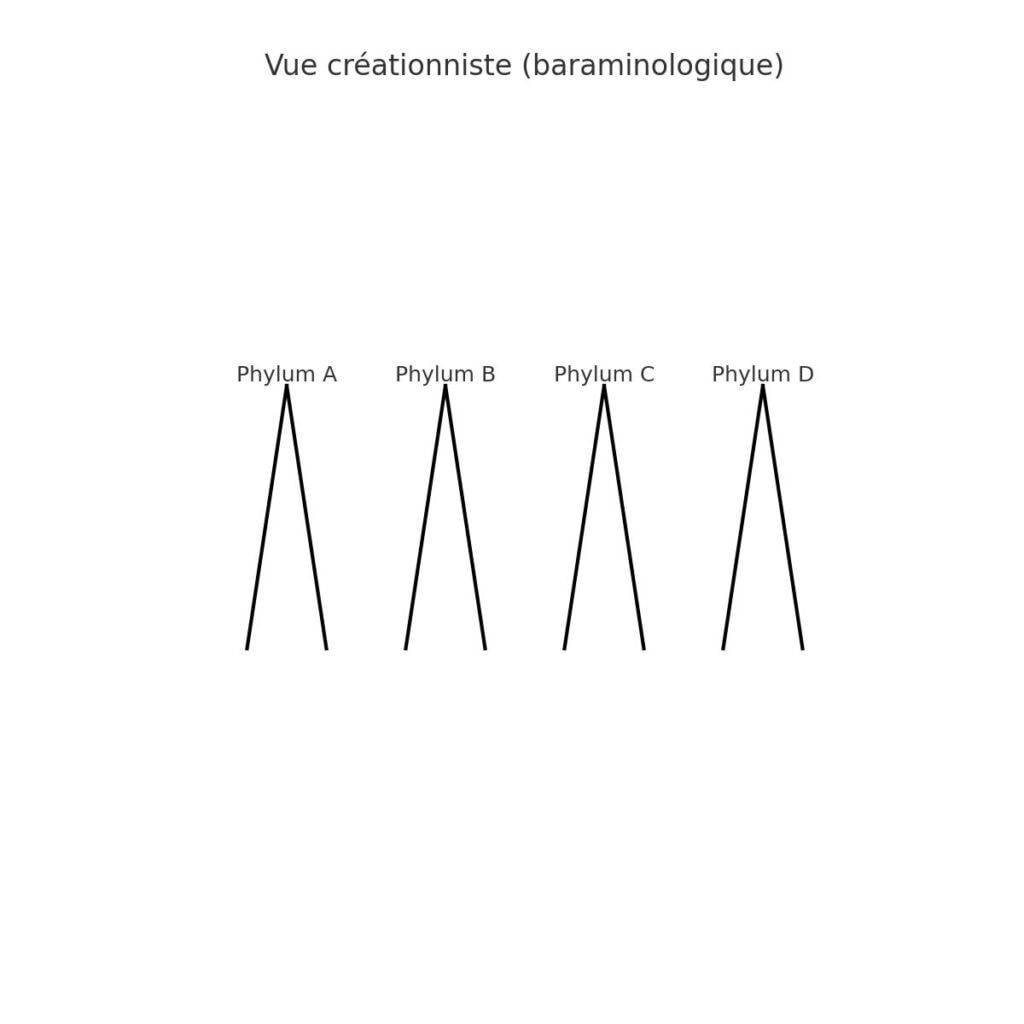
Les grands groupes taxonomiques (phyla) apparaissent en premier dans le registre fossile (ou du moins plus bas dans la colonne), suivis de niveaux inférieurs. Cela ne correspond pas à un arbre évolutif progressif partant du bas. Cela suggère plutôt:
- Une apparition soudaine de plans corporels majeurs (création ou explosion d’innovation initiale),
- Suivie d’une tendance descendante (diversification, spécialisation).
C’est un argument fort en faveur d’une création par types originels suivie d’adaptations, comme proposé par de nombreux créationnistes.
| Élément | Modèle évolutionniste | Modèle créationniste |
|---|---|---|
| Rangs taxonomiques | Représentent des niveaux d’ascendance commune | Représentent des catégories de forme/fonction |
| Origine des grands groupes | Phylogénie ascendante (unicellule → complexité) | Création directe de types distincts |
| « Espèce » | Unité de sélection évolutive | Peut résulter de diversification post-création |
| Baramin | Non reconnu | Unité fondamentale de la Création, souvent ≈ famille, genre, clade |
| Apparition dans les fossiles | Progressive, ascendante | Soudaine, complète, stable |
Il est important de se déformater du récit évolutif pour comprendre autrement le registre fossile. Toutes les « familles » d’organismes ont vécu en même temps, c’est juste qu’ils n’ont pas été emportés en même temps lors des événements séquentiels du Déluge. La progression de la mer vers la terre ne représente pas une évolution mer-terre mais la montée progressive des eaux. Le déluge a commencé dans les océans avec « l’éclatement des sources du grand abîme » de Genèse 7:11. L’eau est ensuite montée progressivement sur les continents jusqu’à finalement tout recouvrir.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!