
Réponse créationniste à la vidéo « les preuves de l’évolution » de Passe-Science – sur les preuves paléontologiques (4)
Dans ce quatrième article, nous continuons notre révision de la vidéo « Les preuves de l’évolution » de la chaîne YouTube « Passe-Science ». Du premier au troisième article nous avions traité la première preuve qui était « les preuves anatomiques« , la deuxième preuve qui était « les preuves développementales » et la troisième preuve qui était « les preuves génétiques« . J’aborde ici le quatrième type de preuve: « les preuves paléontologiques ».
4. preuves paléontologiques
Cabaret commence en disant:
« Tous ces fossiles sont des preuves directement observables que les formes de vie sur terre ont changé au fil du temps. Leur nombre et la richesse des données qui s’y rattachent permet dans certains cas de faire des statistiques.«
Cette affirmation est contestable. Il n’y a aucune preuve que les formes de vie sur terre ont radicalement changé au fil du temps. On observe des variations minimes au sein des genres ou familles mais rien qui ne transforme un organisme radicalement en une autre forme fondamentale.
Pour démontrer que les formes de vie ont changé au fil du temps, il aurait fallu montrer les « innombrables » transitions que Darwin attendait et qui n’ont jamais vu le jour malgré un registre fossile plutôt complet (voir Michael Denton « Evolution: A Theory in Crisis en 1985 » puis « Nature’s Destiny et Evolution: Still a Theory in Crisis en 2016 »).

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
Les formes de vie retrouvée sont abouties, sans relation ancestrale ou intermédiaire pouvant prouver qu’elles ont changé. Cette discontinuité s’insère très bien dans le modèle baraminologique créationniste où Dieu a créé des genres différents à l’origine (par exemple un couple originel d’éléphants pouvant varier en éléphant d’Asie, d’Afrique et mammouth). Cette « forêt de la vie » biblique correspond à la « forêt de fossiles » observée en paléontologie.
On peut aussi mentionner les fossiles vivants, ou le taxon de Lazare. Il s’agit d’espèces connues à l’état fossile (parfois supposée disparue depuis des millions d’années) et qui existe encore aujourd’hui, presqu’inchangée. Il existe plusieurs exemples connus:
- Les cœlacanthes qui sont censés avoir disparu il y a 65 millions d’années, mais découvert vivant en 1938.
- Les ginkgo biloba, nautiles, limules, séquoias, fourmis fossiles, crevettes, crocodiles qui sont identiques à leurs homologues modernes, etc.
Ces espèces n’ont pas changé malgré les millions d’années supposées d’évolution. Cela contredit l’idée de mutations constantes et de transformation progressive et l’absence de forme intermédiaire renforce l’idée d’une création telle que décrite dans la Genèse.
L’idée chez les évolutionnistes est que les organismes n’ont existé ou n’existent que durant la période qui correspond aux couches géologiques dans lesquelles ils reposent. Si une espèce n’est pas trouvée dans un niveau géologique, alors il n’a tout simplement pas vécu à ce moment là et il n’a pas côtoyé les autres espèces qui y sont trouvées. Les fossiles vivants mettent en doute cette « hypothèse » et soutiennent le modèle séquentiel du registre géologique dans le Déluge biblique (des mers vers les terres).
Ce n’est pas que le cœlacanthe n’existait pas à l’époque correspondante de certaines couches géologiques où il n’est pas trouvé mais qu’il ne se trouvait pas sur les écosystèmes dévastés à ce moment là. Le Déluge, de manière générale, a emporté les organismes écosystème par écosystème. Les tsunamis, le volcanisme, le reflux des eaux ont créé des mélanges terrestres/marins mais nous pouvons observer un cadre général dans le registre fossile allant des mers vers les terres (invertébrés marins, poissons, amphibiens, reptiles, mammifères, oiseaux….).
Ce sujet est amplement détaillé dans les articles suivants:
- L’Évolution et l’épique quête des fossiles intermédiaires et des ancêtres évolutionnaires
- De l’Arbre de la Vie de Darwin à la Forêt de la Vie de Moïse : Deux visions de l’histoire du vivant


Cabaret poursuit en disant:
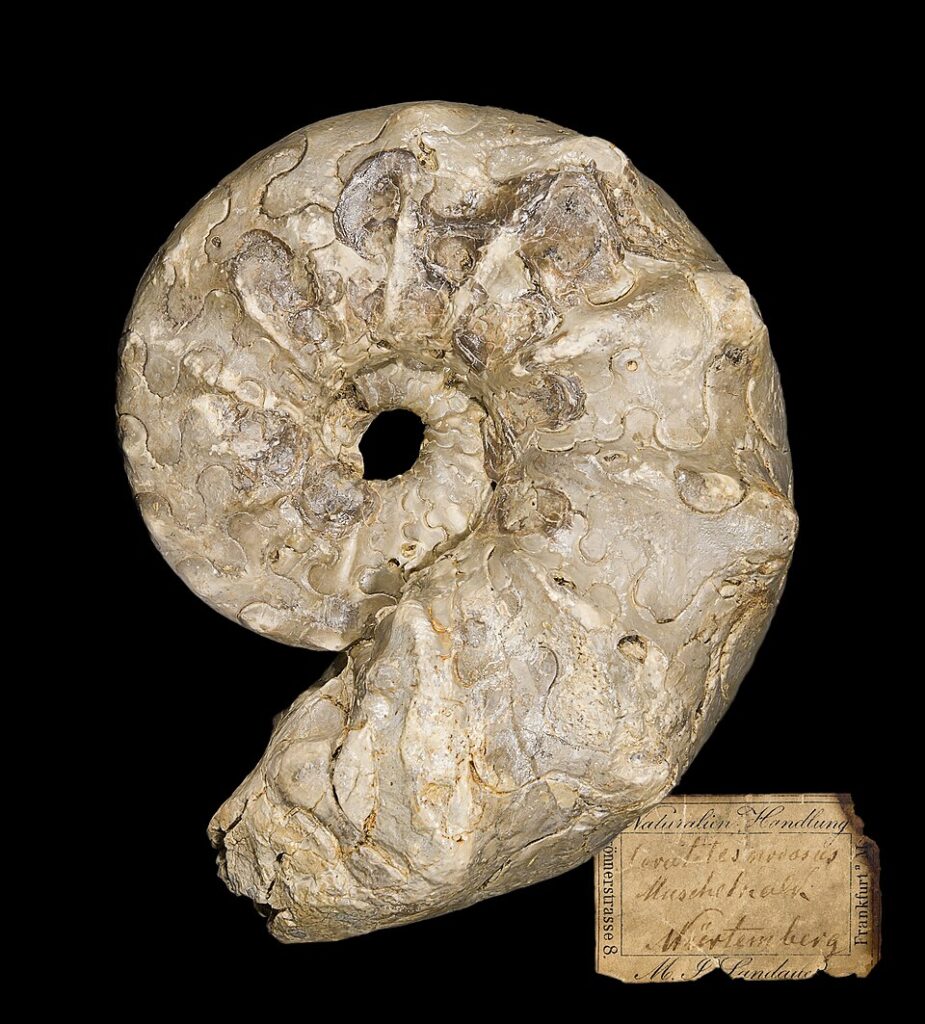
« On va s’intéresser à ces quatre ordres d’animaux (Orthocerida, Goniatitida, Ceratitida, Ammonitida). La différence entre les deux premiers est évidente, elle l’est beaucoup moins sur cette illustration pour les trois suivants. Mais ces organismes sont très différents. Si on regarde les fossiles associés, on peut constater des différences au niveau des sutures entre compartiments. Initialement très simple puis progressivement plus complexe. Au niveau de la structure interne c’est encore plus flagrant. Vous voyez ici la structure interne fractale d’une ammonite. Dans ces fossiles on peut donc suivre l’évolution d’animaux très distincts depuis des formes initialement massives et rudimentaires jusqu’à l’apparition progressive de structures plus légères et optimisées. Ce qui s’est accompagné via l’optimisation du poids de l’augmentation de la taille moyenne de ces animaux.
Certains membres de notre 4ème famille des ammonites peuvent atteindre plusieurs mètres. Si j’ai choisi ces 4 représentants fossiles (Orthocerida, Goniatitida, Ceratitida, Ammonitida), c’est parce que leur abondance permet de tracer des courbes et les voici. En plus de l’évolution morphologique qu’on a pu constater directement sur les fossiles, on voit nettement sur ces courbes que chaque ordre connaît son âge d’or avant le suivant. L’explosion des cératites ici fait suite à l’extinction permien/trias ayant fait disparaître concurrence et prédateur. L’ensemble de ces données paléontologiques témoigne assez clairement de lien évolutif entre ces populations animales. Ceci nous donne un aperçu de la précision dans la reconstruction du passé qu’il est possible d’atteindre sur la base de données paléontologiques. Cette précision est la conséquence de l’utilisation de non pas une mais d’une bonne vingtaine de méthodes de datation très variées comme l’étude de couche au sein des sols ou des méthodes basées sur la désintégration d’isotopes radioactifs. Un large de panel de méthode de datation venant se confirmer et se compléter. »
Les données montrent des variations au sein d’un même groupe, aucune de ces formes ne démontre un changement de genre ou une macroévolution.
L’interprétation uniformitariste des couches et des fossiles ignore l’alternative catastrophiste/créationniste (comme le Déluge mondial), qui peut expliquer à la fois l’abondance et la distribution ordonnée des fossiles.
Avec les ammonoïdes on observe des variations et non des transitions. Les orthocéridés, goniatites, cératites, ammonites sont tous des céphalopodes marins à coquille. Ils diffèrent par la complexité des sutures (jonctions internes entre loges de la coquille) et la taille moyenne et le degré de courbure. Ces différences sont modulaires et répétitives (formes fractales dans les sutures). Elles n’impliquent aucun nouvel organe, aucun nouveau plan corporel. Cela peut être vue comme une diversification intra-baramin (variabilité dans le même genre créé).
C’est comme si on montrait la diversité des cornes chez les bovins ou des plumes chez les oiseaux, c’est effectivement impressionnant, mais ce n’est pas un changement de « genre ».
Les évolutionnistes ont une lecture chronologique où chaque groupe succède au précédent mais une autre lecture est possible. Les couches géologiques ne sont pas toujours du temps accumulé, mais souvent des dépôts catastrophiques rapides (cf. volcanisme, turbidités, Déluge mondial). Les “âges d’or” des groupes fossiles pourraient refléter des zones écologiques englouties successivement pendant la catastrophe (écologie zonale) et non une succession temporelle de millions d’années.
Les ammonites sont proliférantes au Déluge, ensevelies massivement, souvent utilisées comme fossiles-index, mais cela prouve surtout une distribution stratigraphique ordonnée par habitat et pas nécessairement par évolution.
Les évolutionnistes voient la « progression graduelle » d’un design simple vers un design optimisé, alors que les formes simples et complexes coexistent dans le registre fossile. Les sutures fractales ne sont pas une innovation évolutive, mais une variation géométrique répétée dans un plan de coquille déjà existant. La complexité ne suit pas une vraie « échelle ascendante », mais des alternances de simplification et de complexification suivant les lignées.
Cabaret évoque la « vingtaine de méthodes convergentes ». En fait, toutes reposent sur des présupposés uniformitaristes (constance des taux de désintégration, systèmes fermés, absence d’événements catastrophiques majeurs). Les méthodes de corrélation stratigraphique (fossiles-index, couches) sont circulaires: on date la couche par les fossiles, puis on « prouve » l’évolution grâce à la chronologie des fossiles. Les isotopes eux-mêmes donnent souvent des résultats discordants ou nécessitent des “corrections” pour coller au scénario.
Plusieurs articles sur le site explorent en détail les problèmes de la datation radiométrique:
- Datation Radiométrique – Age des Roches, Fossiles, de la Terre et de l’Univers – Créationnisme
- Les Fossiles d’Homo Sapiens à Djebel Irhoud ont-ils vraiment 300 000 ans?
- Le Carbone 14 dans les Diamants, le Charbon, les Fossiles et les Dinosaures
On présente les ammonites comme un bel exemple d’évolution progressive avec des sutures de plus en plus complexes, des tailles croissantes et des groupes successifs. En réalité, ce sont toujours des ammonoïdes, la même catégorie de céphalopodes, avec des variations de coquille. Les formes simples et complexes coexistent, et leur abondance massive s’explique très bien par un dépôt catastrophique, comme celui du Déluge. Les datations invoquées ne sont pas neutres, elles reposent sur des présupposés uniformitaristes. Ce que l’on observe n’est pas une transition macroévolutive, mais une diversité dans un même groupe d’organismes, enseveli massivement lors d’une catastrophe.12
Cabaret poursuit en proposant les candidats Archeopteryx et Tiktaalik comme fossiles intermédiaires ou transitionnels. Ces derniers malheureusement ne s’insèrent pas bien dans le récit évolutif. Dans le premier cas, il existe des traces d’oiseaux dans des couches inférieurs à Archaeopteryx, dans le deuxième des traces de tétrapodes avant Tiktaalik. En gros cela voudrait dire que les petits-fils apparaissent avant les grands-parents dans l’histoire. En fait Cabaret a dû recevoir des commentaires lui indiquant le problème puisqu’il a ajouté dans la section commentaire sous la vidéo:
« Si l’on inclut des publications spécifiques plus récentes, on peut trouver des fossiles de tétrapodes qui chevauchent ou précèdent ceux de Tiktaalik. C’est attendu, car lorsqu’on découvre des traces du passé, on n’a pratiquement aucune chance de tomber sur un ancêtre direct. Ainsi, Tiktaalik n’est pas véritablement l’ancêtre direct des tétrapodes, mais un descendant de cet ancêtre commun, dont il reste très proche. L’évolution de la courbe du nombre d’occurrences de fossiles de tétrapodes est assez parlante. »
Comme l’indique Michael Denton ou d’autres, on ne peut pas accuser le registre fossile d’être incomplet. Il y a des trilliards de fossiles, il n’y a aucune raison de trouver les formes terminales et non les formes intermédiaires s’ils ont existé. Cela révèle qu’il y a un problème avec cette théorie. Le sujet est amplement couvert dans les articles suivants (en particulier pourquoi on a cette caractéristique récurrente de traces qui apparaissent avant les parties dures des organismes):
- Le Registre Fossile Interprété par les Créationnistes
- L’Évolution et l’épique quête des fossiles intermédiaires et des ancêtres évolutionnaires
- Comment se forment les fossiles ? Lentement ? Rapidement ?
Malgré tout, de nombreux évolutionnistes persistent à argumenter des fossiles transitionnels, comme ceux qui auraient mené les thérapsides a évolué en mammifères ou encore Archæoptéryx. Ce dernier est intéressant, il dispose de caractéristiques mosaïques avec les oiseaux et les reptiles mais c’est la même chose des oiseaux et mammifères modernes. Ça n’indique pas nécessairement une relation évolutive.
Les plumes d’Archæoptéryx ne constituent pas une transition entre les écailles et les plumes, car ses plumes sont abouties et optimisées. On n’y perçoit pas un stage intermédiaire. Les évolutionnistes peuvent argumenter l’évolution rapide, mais si nous n’avons que des écailles optimisées d’un côté et des plumes entièrement formées de l’autre, il est difficile de voir ce qu’il y a d’intermédiaire dans Archæoptéryx.
Il s’insère très bien dans un groupe baraminologique créationniste. Il pouvait visiblement accomplir des vols courts et se déplacer avec agilité dans les arbres.
Une chauve-souris n’est pas une forme de transition entre l’oiseau et le mammifère terrestre, et un ornithorynque n’est une forme de transition entre le canard et le mammifère, même s’il présente certaines caractéristiques des deux. Si donc les mosaïques de caractères de la chauve-souris et de l’ornithorynque ne nécessitent pas de relation évolutive, pourquoi la mosaïque d’Archæoptéryx devrait absolument refléter un intermédiaire entre reptile et oiseau.
Même l’évolutionniste Barbare Stahl déclare:
« Puisqu’Archaeopteryx occupe une position isolée dans le registre fossile, il est impossible de dire si cet animal a donné naissance à des formes volantes plus avancées ou s’il ne représentait qu’une branche latérale par rapport à la lignée principale. »
Elle a ajouté dans une édition ultérieur (Dover 1985):
« La découverte de véritables fossiles d’oiseaux datant du Crétacé inférieur n’a fait que renforcer l’argument selon lequel le célèbre Archaeopteryx à plumes pourrait être une branche archaïque dérivée de la lignée avienne ancestrale. »
Stahl, B.J., p. Vertebrate History: Problems in Evolution (McGraw–Hill 1973).
Les évolutionnistes nient la présence de traces d’oiseaux bien avant Archaeoteryx, mais cela se comprend, les admettre mettrait fin au peu d’arguments restants sur la transition reptile/oiseau.
Le Dr Jake Hebert fait bien le point sur ces traces d’oiseau et la réaction évolutionniste.3 Il rapporte par exemple que le paléontologue amateur et évolutionniste Jerry MacDonald avait mené des recherches paléontologiques novatrices au Nouveau-Mexique4. Parmi ses découvertes, il avait trouvé des empreintes dans des roches du Jurassique qui ressemblaient étrangement à des traces d’ours, d’oiseaux et de singes.
« Les empreintes fossiles que MacDonald a collectées incluent un certain nombre de ce que les paléontologues aiment appeler des « problematica ».
Sur une piste, par exemple, une créature à trois doigts a apparemment fait quelques pas, puis a disparu — comme si elle s’était envolée. « Nous ne connaissons aucun animal à trois doigts au Permien [298.9 Ma – 251.902 Ma selon le référentiel évolutif] », souligne MacDonald. « Et il n’est pas censé y avoir d’oiseaux. »
Il possède plusieurs pistes où des créatures semblent marcher sur leurs pattes arrière, d’autres qui paraissent presque simiennes [semblables à des singes].
Sur une paire de tablettes de siltstone, je remarque des empreintes inhabituellement grandes, profondes et inquiétantes, chacune avec cinq marques de doigts arqués, comme des griffes. Je fais remarquer qu’elles ressemblent exactement à des traces d’ours. « Oui », dit MacDonald à contrecœur, « c’est bien ce qu’elles semblent être. » Les scientifiques s’accordent à dire que les mammifères ont évolué bien après la période permienne, et pourtant ces empreintes sont clairement permiennes.«
Stewart, D. 1992. Petrified Footprints: A Puzzling Parade of Permian Beasts. Smithsonian. 23 (4): 70–79.
De nombreux évolutionnistes (mais pas tous) savent que le registre fossile n’est pas l’endroit privilégié pour argumenter l’évolution. Il correspond beaucoup plus aux attentes bruts du modèle biblique.
«Les espèces fossiles demeurent inchangées au cours de leur histoire, et le registre fossile ne contient pas un seul exemple transitionnel significatif.»
David S Woodruff, « Evolution : The paleobiological views », Science, vol 208, 16 may 1980, p 716
« Aucun véritable évolutionniste n’utilise les archives fossiles comme preuve de l’évolution plutôt que de la création.«
M. Ridley, 1981, Who doubts evolution? New Scientist, 90:832
De son côté le zoologiste français Pierre Grassé déclarait:
« Les naturalistes doivent se rappeler que le processus de l’évolution n’est révélé que par les formes fossiles. Une connaissance de la paléontologie est donc un préalable ; seule la paléontologie peut leur fournir les preuves de l’évolution et en révéler le cours ou les mécanismes. Ni l’examen des êtres actuels, ni l’imagination, ni les théories ne peuvent se substituer aux documents paléontologiques. »
Evolution of Living Organisms – Academic Press, New York, 1977, p. 4
D’aucuns pourraient dire que la théorie de l’évolution est réfutée et la prédiction de Darwin sur les innombrables fossiles transitionnels non réalisée, mais il y a quelque chose de religieux dans l’évolution et c’est pourquoi l’argument évolutionniste est résolument orienté vers la génétique, comme nous l’avions vu pour les trois premières preuves et la cinquième que nous allons réviser dans l’article suivant.
Commentaire personnel
Dernier commentaire très personnel, et sans lien direct avec la vidéo de Passe-Science, sur l’ancestro-manie où la recherche effrénée de preuves d’ascendance commune chez les évolutionnistes.
Il y a, me semble-t-il, dans une large partie de la recherche scientifique contemporaine une véritable obsession pour l’ascendance commune – une « ancestro-manie » qui dépasse parfois le cadre de la rigueur scientifique pour devenir une quête idéologique. Cette frénésie interprétative, typique de la pensée évolutionniste moderne, conduit trop souvent à forcer les données, à interpréter les faits selon un schéma préétabli, à surinterpréter des ossements, et à ignorer des lectures alternatives pourtant raisonnables.
Prenons les rétrovirus endogènes (ERV). Loin d’être des vestiges muets d’anciennes infections virales, ils révèlent aujourd’hui des fonctions biologiques cruciales dans la régulation des gènes, le développement embryonnaire, le placenta, ou l’immunité. Et pourtant, l’interprétation dominante reste obstinément figée : ce sont les preuves d’une infection ancestrale. Quand on découvre qu’un ERV est fonctionnel, il devient « coopté » par hasard ; s’il est partagé entre espèces, c’est « preuve d’un ancêtre commun » ; s’il ne l’est pas, on invoque une perte ou une insertion indépendante. Autrement dit : tout confirme le récit évolutionniste, même ce qui le contredit.
Même chose pour la fusion chromosomique supposée à l’origine du chromosome 2 humain. L’histoire est belle sur le papier, mais elle repose sur des séquences ambiguës, des sites de fusion dégénérés, et une interprétation qui suppose la conclusion à prouver. Que ces régions puissent être des séquences normales, fonctionnelles, ou issues d’un design particulier n’entre même pas dans le cadre de la discussion.
Et que dire des structures dites « vestigiales » ? Elles sont, dans bien des cas, parfaitement fonctionnelles, comme l’appendice ou le coccyx. Mais peu importe : elles sont proclamées “restes” d’une époque antérieure. L’évolution n’explique pas leur présence ; elle l’assume.
Quant aux fossiles « transitionnels », on en fait des vedettes médiatiques sans preuve réelle de transition biologique ou génétique. L’évolution est ici moins un modèle explicatif qu’un prisme interprétatif, un narratif dominant auquel tout doit se conformer.
Et puis, bien sûr, il y a l’argument du « mauvais design » : si une structure est imparfaite à nos yeux, alors c’est la preuve qu’elle n’a pas été conçue. C’est une logique curieuse : on évalue le design non à sa fonction réelle, mais à notre compréhension partielle, biaisée, ou esthétique. Cette critique n’est pas scientifique — elle est théologique.
Enfin, l’évolution convergente, ce concept « sauveur » qui explique les similitudes quand l’ancêtre commun ne suffit plus, trahit elle-même l’incapacité du modèle évolutionniste à expliquer de manière cohérente la récurrence de certains motifs biologiques complexes. Pourquoi les mêmes structures apparaissent-elles indépendamment dans des lignées supposées éloignées ? Réponse : « par convergence »…
Cette ancestro-manie, c’est finalement le symptôme d’une science dominée par une métaphysique naturaliste. Toute la biologie est interprétée à travers la lentille de la descendance commune, et tout ce qui ne rentre pas dans ce cadre est soit ignoré, soit tordu jusqu’à ce qu’il s’y plie.
Mais d’un point de vue créationniste, on peut — et on doit — lire autrement ces données. Les ERV, les structures répétées, les similitudes entre espèces, les pseudo-vestiges, les « mauvais designs », tout cela peut s’expliquer par un plan commun, une architecture intelligente, une diversité intégrée. Ce ne sont pas les restes d’une histoire sans but, ce sont les indices d’un dessein rationnel et ordonné.
Mais au-delà de l’obsession pour la descendance, ce qui me frappe encore davantage, c’est le double standard méthodologique et intellectuel que l’on rencontre dans ce débat. Les créationnistes sont souvent accusés d’être trop “littéralistes”, d’avoir une lecture “figée” ou “pré-scientifique” du texte biblique. Pourtant, leur lecture — qu’on l’accepte ou qu’on la conteste — est cohérente, assumée et historiquement enracinée, ancrée dans une herméneutique naturelle du texte, telle qu’elle était comprise par Jésus, les prophètes, et l’Église historique pendant des siècles.
Mais ce que beaucoup oublient, c’est que les évolutionnistes ont eux aussi leur texte fondateur : la descendance universelle à partir d’un ancêtre commun, dogme central du darwinisme. Et ils y sont souvent plus attachés que bien des chrétiens à leur propre Bible.
Ce n’est pas une simple métaphore : leur lecture de la nature passe par une interprétation a priori du monde biologique, tout comme un théologien pourrait le faire d’un texte sacré. Et ce zèle quasi-religieux les a parfois conduits à défendre, justifier ou taire des fraudes — pour préserver leur “paradigme”.
- L’homme de Piltdown (1912) : un crâne truqué (mâchoire de singe + crâne humain) accepté pendant 40 ans comme “preuve de transition” par la communauté scientifique.
- L’homme du Nebraska (1922) : reconstruit à partir d’une dent de cochon — utilisé dans le procès Scopes.
- Archaeoraptor (1999) : un faux fossile chinois, assemblé avec des pièces collées de différentes espèces — publié dans National Geographic.
- Les embryons d’Haeckel (1860s) : exagérés ou totalement falsifiés pour montrer des similarités fictives entre embryons — utilisés pendant des décennies dans les manuels malgré des dénonciations précoces.
- Les “fossiles chinois truqués” : comme l’a montré Carl Werner, des centaines de fossiles auraient été retouchés, reconstitués ou même fabriqués dans certains circuits de marché noir, souvent pour confirmer un récit évolutif déjà admis.
Dans chacun de ces cas, la soif de confirmer la théorie a supplanté la prudence scientifique.
L’homme de Néandertal
Un autre exemple frappant de cette « ancestro-manie » idéologique, c’est la manière dont l’homme de Néandertal a été représenté pendant des décennies.

Alors que les preuves morphologiques, génétiques, et archéologiques montrent aujourd’hui clairement que Néandertal était pleinement humain — avec un cerveau souvent plus volumineux que celui de l’homme moderne, des outils, des rites funéraires, de la musique (flûte de Divje Babe), et un langage articulé — les premières reconstitutions le montraient comme un être simiesque, voûté, hirsute, à demi-bête.
Cette image caricaturale, issue de dessins de la fin du XIXe siècle (notamment ceux inspirés par le travail de Marcellin Boule), a profondément marqué l’imaginaire collectif. Elle a contribué à enraciner dans l’esprit du public l’idée que l’homme descend du singe, au mépris de ce que les données réelles indiquaient déjà.
Et même lorsque la science a corrigé ces erreurs (ex : Boule avait mal interprété un squelette arthritique), les manuels, musées et films ont continué pendant des décennies à diffuser l’image du “néandertalien-singe”, parfois jusqu’à aujourd’hui.


 Neanderthal-Museum, Mettmann
Neanderthal-Museum, Mettmann
C’est un exemple parmi d’autres où la vision du monde darwinienne a devancé les données, jusqu’à les façonner visuellement, émotionnellement, culturellement. Une science du récit, plus qu’une science des faits.
On oublie que dans l’esprit du public, ce ne sont pas les publications scientifiques qui forgent les convictions, mais les illustrations, les diaporamas de musées, les films éducatifs, les animations 3D. Et ce sont souvent là que le biais évolutionniste s’exprime le plus librement.
Alors que les fossiles humains anciens sont souvent fragmentaires, les reconstitutions les transforment en portraits détaillés, peau nue, regard torve, posture animale, comme s’il fallait absolument visualiser la transition entre “bête” et “homme”. Mais ces images ne sont pas des données, ce sont des interprétations, souvent guidées par une intention narrative.
Pezosiren portelli
Un autre cas emblématique de cette obsession pour les formes de transition est celui du Pezosiren portelli, parfois présenté comme un « chaînon manquant » entre les mammifères terrestres et les siréniens (lamantins, dugongs). Ce fossile, découvert en Jamaïque, a été immédiatement interprété par le paléontologue évolutionniste Daryl Domning comme une « vache de mer à pattes » – une transition entre quadrupède terrestre et mammifère marin.
Mais une lecture critique révèle plusieurs problèmes troublants:
- Le squelette reconstruit est composite, constitué d’éléments issus de plusieurs fossiles éparpillés dans des strates contenant aussi bien des os de rhinocéros, de crocodiles, que de siréniens marins typiques.
- La présence simultanée d’ossements de créatures terrestres et marines dans la même couche sédimentaire suggère plutôt un dépôt catastrophique mélangé, et non une preuve de transition évolutive.
- La fameuse reconstruction repose sur des interprétations spéculatives : le crâne n’est quasiment pas illustré, et de nombreux éléments essentiels sont absents ou non clairement attribuables à un individu unique.
Andrew Lamb déclare:5
« Les os rassemblés par Daryl Domning pour construire ce qu’il a nommé Pezosiren portelli provenaient de trois lits osseux distincts au sein du membre géologique de Guys Hill, épais de cinq mètres, en Jamaïque. (Notons que, dans la pensée évolutionniste, ces cinq mètres représenteraient plusieurs millions d’années de sédimentation accumulée.)
Certains des os ont été trouvés isolés et d’autres sous forme de squelettes partiels. La strate en question a livré plusieurs centaines d’os, dont ceux d’un rhinocéros (Hyrachyrus), d’un lézard, d’un crocodile (Charactosuchus kugleri), d’une tortue, de dugongs (« restes abondants de siréniens »), et peut-être d’un primate, ainsi qu’un grand nombre de fossiles marins invertébrés (mollusques, etc.).
Il me semble donc très probable qu’en construisant sa “vache de mer à pattes”, Domning ait combiné des os provenant de différentes sortes de créatures.
Domning a omis d’inclure dans son article de Nature des photos de la plupart des os, y compris du crâne ou de fragments de crâne (à l’exception d’une partie de la mâchoire droite). »
Tout cela illustre un biais persistant: le désir de trouver coûte que coûte des ancêtres là où un regard plus neutre verrait simplement un assemblage artificiel.
Conclusion
Cette foi dans la théorie est rarement remise en question. Les faits ne sont pas évalués avec neutralité : ils sont organisés, recyclés, parfois modifiés, pour s’aligner avec le récit de l’ascendance commune. Et tout comme on accuse les créationnistes de voir « ce qu’ils veulent voir », on devrait aussi se demander combien de chercheurs évolutionnistes sont eux aussi prisonniers de ce qu’ils espèrent confirmer.
Peut-être faudrait-il cesser de se moquer des « lecteurs littéralistes de la Genèse » pour regarder avec honnêteté ceux qui lisent la nature comme s’il s’agissait d’un texte déjà interprété, déjà canonisé par Darwin.
La vraie science ne devrait pas chercher à confirmer des dogmes, mais à suivre les données là où elles mènent. Et si l’on fait cela honnêtement, alors peut-être découvrira-t-on que le récit créationniste, loin d’être une fuite dans la foi, est au contraire une grille de lecture cohérente, fertile et sérieuse.
Dans l’article suivant, nous étudierons le cinquième preuve apportée dans la vidéo sur les preuves de l’évolution: les observations directes.
- https://www.icr.org/content/mind-blowing-marine-ammonite-tree-resin.
- https://www.icr.org/content/trilobites-sudden-appearance-and-rapid-burial.
- https://www.icr.org/article/bird-like-tracks-before-birds.
- MacDonald, J. 1994. Earth’s First Steps: Tracking Life Before the Dinosaurs. Johnson Printing. Boulder, CO.
- https://creation.com/ancient-mutant-jamaican-sea-cows.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!