

Les Preuves de la Création et du Déluge: Paléontologie (3)
J’ai abordé dans le premier article de cette série les hypothèses du modèle créationniste et comment j’articule ma recherche sur cette enquête des origines. Dans le deuxième article, j’avais exploré « les preuves en biologie« . Je traite dans ce troisième article de la paléontologie qui est peut-être le domaine le plus concret pour les néophytes.
- Baraminologie / Paléontologie
- Perspective descendante du registre fossile
- Les traces de pas
- Rapidité de la fossilisation
- Les fossiles vivants
- Le carbone 14
- Les biomatériaux
- La progression mer-terre
- La disparition des dinosaures
- Légendes de déluge
- Pictogramme chinois
- Echos de la tour de Babel
- Les langues
- Récits de dragons
- Ere glaciaire
- Biogéographie
- Perte de technologies (chasseurs-cueilleurs, agriculteurs)
- Défenses évolutionnistes
Baraminologie / Paléontologie
Les concepts de baraminologie (genres créés) et de paléontologie sont liés. La baraminologie est l’étude des genres créés dans Genèse, l’idée est que Dieu a créé des groupes d’organismes à l’origine, cette logique doit être observé dans le registre fossile si le créationnisme est vrai. Nous devrions observer une forêt de la vie (plutôt qu’un arbre).
L’espèce biblique n’est pas l’espèce moderne, le contour de l’espèce biblique est déterminé principalement par le continuum reproductif. Par exemple, âne, zèbre et cheval sont des espèces différentes, mais bibliquement elles sont une espèce car ils peuvent se reproduire ensemble (on utilise alors le mot « genre » plutôt qu’espèce).
En baraminologie, tous les équidés (Equus caballus, E. asinus, E. zebra, E. quagga, etc.) sont vus comme faisant partie du même « genre créé » (holobaramin). Après le Déluge, un couple d’équidés fondateurs aurait donné naissance à l’ensemble des chevaux, ânes et zèbres actuels par spéciation rapide. La possibilité de produire des hybrides (même stériles) est un argument fort de continuité génétique au sein d’un baramin.

Inscrivez vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
Le continuum n’a pas nécessairement à être complet, parce que les continuums morphologique et comportemental peuvent aider à retrouver un baramin. Par exemple chat et tigre ne peuvent pas se reproduire alors que la plupart des créationnistes les classeraient dans le même genre, mais nous savons qu’un chat est en quelque sorte un petit tigre et un tigre un gros chat.
Tous les félins (du tigre au chat domestique) peuvent être reliés par chaîne d’hybrides documentés.
Tigre → Lion → Léopard → Puma → Ocelot → Margay → Chat domestique → Chat à pieds noirs.
Ainsi, même si le chat domestique et le tigre ne s’hybrident pas directement (notamment à cause de la taille, du poids et des comportements), ils sont reliés via des ponts reproductifs ou morphologiques (par exemple via serval –> léopard –> lion/tigre). Le tout s’inscrit dans un cadre génétique commun et une morphologie très conservée au sein des félins.
La baraminologie créationniste prédit donc une « forêt de la vie suspendue », sans lien généalogique entre grands groupes d’organismes. Il y a des délimitations claires. Les groupes peuvent varier en leur sein et produire une diversité de félins, de canins, d’ursidés etc… mais on ne traverse pas les genres. Aucune observation dans le présent ou dans le passé (registre fossile) ne démontre le contraire.
Perspective descendante du registre fossile
Quand on fait une lecture du registre fossile en partant du bas de la colonne géologique, là où apparaissent les animaux, pour finir en haut de la colonne, on constate une direction compatible avec la notion baraminologique (nous parlerons ensuite de comment le déluge explique le dépôt quasi simultanée de toutes les couches, en l’espace d’un an).
La direction observée, taxonomiquement parlant, est qu’on ne part pas du bas de la classification taxonomique, mais d’en haut. Sur environ 35 phylums actuels (selon classification), au moins 20 à 27 apparaissent brusquement dès le Cambrien. Parmi eux:
- Arthropodes (trilobites, marrellomorphes, etc.)
- Mollusques (formes bivalves, gastéropodes primitifs)
- Échinodermes (formes basales comme les stylophores)
- Chordés (ex. Pikaia, apparenté aux céphalocordés ; ancêtres possibles des vertébrés)
- Cnidaires (formes apparentées aux méduses, coraux)
- Annélides (vers segmentés)
- Porifères (éponges)
- Brachiopodes, Priapulides, etc.
Environ 20 phylums sont attestés de façon claire dans les gisements cambrien (Burgess Shale, Chengjiang, Sirius Passet). Certains chercheurs parlent de 85% des plans d’organisation actuels déjà représentés. Des centaines de nouvelles classes apparaissent d’un seul coup. Les trilobites (Arthropodes) seuls comptent plus de 20 ordres diversifiés dès le Cambrien inférieur.
Sepkoski (2002, Compendium of Fossil Marine Animal Genera) recense environ 1 300 genres marins connus pour le Cambrien. D’autres estimations tournent autour de 1 000 à 1 500 genres selon les révisions taxonomiques.
Dans le Précambrien (Édiacarien, 635–541 Ma, selon la chronologie séculière), on trouve des fossiles mous bizarres (les « Édiacariens » comme Dickinsonia, Charniodiscus). Ils n’ont aucun lien évident avec les phylums du Cambrien. Beaucoup semblent représenter une faune éteinte à part, sans descendance claire.
On n’a pas retrouvé de formes intermédiaires graduelles menant aux arthropodes, mollusques, échinodermes ou chordés du Cambrien.
Les genres cambrien (ex. Olenellus, Paradoxides pour les trilobites ; Pikaia pour les chordés) apparaissent déjà typés, avec leurs caractères distinctifs. Les paléontologues admettent qu’il est extrêmement difficile de relier généalogiquement les différents genres cambrien, car les fossiles montrent des morphologies complètes et distinctes, sans étapes intermédiaires.
Même à l’intérieur d’un phylum comme les trilobites, on observe des dizaines de familles bien distinctes dès le Cambrien inférieur. L’idée d’un ancêtre trilobite unique graduel n’est pas documentée dans les fossiles.
- Les phylums apparaissent d’un coup sans ancêtres connus.
- Les genres cambrien sont déjà aussi différenciés que des genres modernes, sans transitions visibles.
Les trilobites pourraient représenter plusieurs kinds distincts, car la disparité morphologique (formes lisses, épineuses, yeux très différents, etc.) pourrait refléter des créations séparées plutôt qu’une seule lignée.
Certains possèdent des yeux composés en calcite avec des propriétés optiques complexes (biomicrolentilles, correction des aberrations). Cela apparaît dès les premiers trilobites cambrien, sans « proto-œil » observé.
Une tendance du simple au complexe n’est pas observée dans le registre fossile. Au contraire, il n’y a qu’à « voir » l’exemple des yeux des trilobites, « les yeux les plus complexes de tous les temps« , qui sont les …. « premiers yeux du registre fossile« … On peut s’imaginer voir clairement dans l’eau avec un champ quasiment de 360°. Certaines espèces (comme les Phacops) possédaient des yeux schizochroaux avec de grandes lentilles individuelles, leur donnant probablement une vision tridimensionnelle (stéréoscopique), en plus du large champ visuel.
On trouve directement les branches principales (20–27 phylums animaux), déjà séparées et bien définies. On ne voit ni le tronc, ni les branches de transition qui devraient précéder. Chaque phylum apparaît sans ancêtre identifié, avec ses propres caractéristiques fondamentales. À l’intérieur même d’un phylum (ex. les trilobites chez les arthropodes), on trouve dès le début une grande diversité (familles et genres distincts).
Cela fait plutôt penser à une forêt de troncs juxtaposés: chaque phylum est comme un arbre déjà adulte, côte à côte, sans racines visibles qui les relieraient. Cela ressemble au modèle biblique des « genres créés » indépendants.
Stephen Jay Gould (paléontologue, Harvard) parlait du Cambrien comme d’une « explosion de disparité ». Au lieu d’une diversification graduelle, on a eu une apparition soudaine de formes très différentes. Gould notait même que la disparité (diversité des plans corporels) était plus grande au Cambrien qu’aujourd’hui.
Simon Conway Morris (Cambridge), spécialiste du Burgess Shale, a reconnu que les archives fossiles donnent plus une apparence de forêt que d’arbre classique (mais il y voit un problème de conservation et de biais fossile).
La prédiction créationniste me paraît satisfaite. Elle postule à la base des groupes d’organismes séparés, donc plusieurs arbres séparés. Dans le registre fossile, on part de grandes catégories, et on varie à l’intérieur de ces catégories.
Concernant les quelques candidats transitionnels (ou intermédiaires structuraux) mis en avant par la concurrence:
- Tiktaalik est-il l’ancêtre des tétrapodes?
- Archæoptéryx est-il l’ancêtre des oiseaux?
- L’Évolution des Baleines est-elle démontrée?
- Les Mammifères ont-ils évolué à partir des Thérapsides (Reptiles Mammaliens)?
- L’Australopithèque est-il l’ancêtre de l’homme?
Les traces de pas
Dans le cadre créationniste et du Déluge biblique, les premiers stades du Déluge produisent de vastes dépôts de boues meubles. Les animaux (ou groupes d’animaux) fuyant les eaux montantes laissent des traces dans ces sédiments encore mous. Ces traces se fossilisent rapidement par recouvrement sédimentaire.
Les organismes sont ensuite emportés par les eaux, ils sont déplacés ou détruits, cela explique pourquoi leurs corps ne se trouvent pas dans les mêmes couches que les traces (les corps sont souvent plus haut). Le tri hydrodynamique sépare les empreintes (restées en place) des cadavres, qui peuvent être déplacés plus loin avant d’être ensevelis.
Les différentes strates ne représentent pas des millions d’années, mais des phases successives d’ensevelissement durant une catastrophe globale rapide. Cela explique pourquoi on observe des traces précoces: elles ont été laissées avant que les organismes aient été submergés et fossilisés.
Dans plusieurs cas, on observe:
- Des traces de trilobites (appelées cruziana) sont trouvées en dessous de leur parties corporelles dans chaque endroit où l’on trouve des fossiles de trilobites,
- Des traces de dinosaures (empreintes de pas, pistes) qui apparaissent dans des couches plus anciennes que les fossiles d’ossements de dinosaures.
- Des traces d’amphibiens tétrapodes (empreintes de pas) au Dévonien, alors que les fossiles des tétrapodes complets apparaissent plus tard.
- Des traces d’oiseaux (empreintes ressemblant à celles d’oiseaux modernes) dans le Trias supérieur.
- Même des cas controversés de traces humaines signalées dans des niveaux plus profonds que ne le prévoit le modèle évolutionniste (l’étude Gierliński et al. de 2017 décrit des empreintes « humanoïdes » en Crète, datées séculairement de 5,7 Ma, avec une morphologie très proche de l’homme moderne).
Ce modèle est connu sous le nom BEDS (Bioturbation, Erosion, Deposition, Sorting). Il explique plutôt bien les traces de pas (pourtant difficiles à fossiliser) qui apparaissent bien en dessous les parties dures des organismes (beaucoup plus facile à fossiliser).
Rapidité de la fossilisation
La fossilisation est un processus rare et exceptionnel. En principe la nature est très forte pour recycler les déchets organiques de sorte que nous ne croulons pas sous les déchets. Il est dès lors étonnant de constater que la fossilisation est un modèle dominant dans les couches sédimentaires.
Le modèle créationniste attribue cela naturellement au déluge de Noé, qui a fourni les eaux et les sédiments pour faire ce travail et empêcher ce « recyclage naturel ». Sans un événement exceptionnel de recouvrement, les carcasses sont amenées à disparaître en l’espace de quelques jours à quelques semaines pour les tissus mous (chair, peau, organes) et en quelques années pour les parties dures (dents, os). On lit sur la page wikipedia « Skeletonization »1:
« Après la squelettisation, si les charognards ne détruisent pas ou n’emportent pas les os, les acides présents dans de nombreux sols fertiles mettent environ 20 ans à dissoudre complètement le squelette de mammifères de taille moyenne à grande, comme les humains, ne laissant aucune trace de l’organisme. »
La fossilisation n’est pas un processus lent mais rapide. Les carcasses, si elles ne sont pas recouvertes immédiatement, disparaissent complètement en l’espace de quelques mois à quelques années. Plusieurs études l’ont documenté 2345.
Les expériences montrent même que dans un milieu aqueux pauvre en oxygène et sans prédateur, les carcasses se désarticulent en l’espace d’une semaine6.
« Les animaux ont dû se transformer en pierre très rapidement. Sinon, leurs entrailles et leurs branchies se seraient décomposées et auraient disparu. Il semble que ces poissons se soient en réalité changés en pierre en un clin d’œil. »
Fox, D. May 2016. Surprise! Fossils in a Flash. https://www.snexplores.org/article/surprise-fossils-flash
« Mais le fossile allemand [d’ichtyosaure] est également inhabituel en ce qu’il semble s’être fossilisé très rapidement, préservant des tissus mous avant qu’ils ne se décomposent. »
– Johan Lindgren, Université de Lund (cité par LePage, M., déc. 2018)
Les fossiles vivants
Un « fossile vivant » est un organisme retrouvé à la fois dans les couches fossiles dites « anciennes » et encore vivant aujourd’hui, sans pourtant qu’il apparaisse tout le long de la colonne géologique, avec très peu ou pas de changements morphologiques (ex. cœlacanthe, limule, ginkgo, nautiles, crocodiles).
Les fossiles vivants illustrent que les organismes ont été créés selon leur « genre », avec une capacité de variation limitée mais une grande stabilité de fond. Leur présence inchangée du passé à aujourd’hui s’explique bien par une création récente et une catastrophe du Déluge qui a enterré massivement la faune et la flore.
Le célèbre évolutionniste Stephen Jay Gould avait déclaré à leur sujet:
« Le maintien de la stabilité au sein des espèces doit être considéré comme un problème évolutif majeur. »7
Les fossiles vivants prouvent que l’absence de fossiles dans une étape géologique n’est pas une preuve d’absence d’existence au moment où cette étape a été formée. De nombreux organismes sont présents au début de la colonne géologique où sont trouvés les animaux (Cambrien et étape ultérieure), n’apparaissent plus par la suite, mais sont encore vivants aujourd’hui. Leur absence à certaines étapes prouve donc qu’une absence ne signifie pas « inexistence ».
Cela soutient le modèle de zonage écologique créationniste, dans lequel les étapes géologiques représentent des étapes du déluge. La présence de types d’organismes dans différents niveaux géologiques est liée aux habitats qui ont été emportés successivement et qui contenaient une faune et une flore particulière.
Le carbone 14
Le carbone 14 est une méthode datation pouvant dater des objets dans l’échelle des milliers d’années. Elle ne peut plus s’opérer sur des matériaux de plus de 60 000 ans (elle ne fonctionne donc plus théoriquement dans l’échelle des 6 chiffres).
Cette méthode reste dépendante d’hypothèses comme les autres méthodes de datation radiométrique. De manière générale est elle est assez fiable sur les trois derniers millénaires car des méthodes indépendantes archéologiques démontrent qu’elle fonctionne assez bien (recoupement cohérent).
Ces mêmes méthodes indépendantes, à mesure que l’on remonte l’histoire (1400 av.J-C et avant), commence à montrer des décalages importants. Cela a probablement à voir avec les ratios de carbone anciens dans l’atmosphère qui étaient différents de ceux observés par la suite.
Les créatures qui ont vécu et sont mortes dans ces temps très reculés n’ont pas baigné dans les mêmes conditions environnementales avec lesquelles nous sommes familiers aujourd’hui ce qui complique les calculs réalisés avec cette méthode.
Le ratio ¹⁴C/¹²C est la clé du calcul. Il est aujourd’hui mesuré à 1 / 1 000 000 000 000. S’il était par exemple de 1 / 5 000 000 000 000 dans le monde antédiluvien, les résultats obtenus par la méthode ne sont pas les mêmes, bien que l’ordre de grandeur des milliers reste juste (moins de 60 000 ans quoiqu’il arrive).
Aujourd’hui il y a dans l’atmosphère environ 420 ppm de CO₂. Selon les reconstitutions séculières, dans certaines périodes géologiques (Paléozoïque, Mésozoïque), l’atmosphère aurait contenu 2000 à 3000 ppm, soit 5 à 7 fois plus de CO₂ qu’aujourd’hui. Cela signifie qu’il y avait beaucoup plus de molécules de CO₂ dans l’air.
Le carbone naturel est constitué principalement de:
- ¹²C (≈ 98,9 %) -> isotope stable dominant.
- ¹³C (≈ 1,1 %) -> stable, utilisé pour les études paléoclimatiques (δ¹³C).
- ¹⁴C (~1,2 × 10⁻¹² fractionnelle) -> radioactif, produit en continu dans l’atmosphère.
Quand on augmente le CO₂ atmosphérique, on augmente surtout la quantité de ¹²C, et un peu de ¹³C, proportionnellement. La part de ¹⁴C reste minuscule et dépend de la production cosmique (rayons cosmiques + champ magnétique) et pas directement de la concentration totale de CO₂.
Quand on dit qu’il y avait 2000–3000 ppm de CO₂, cela veut dire qu’il y avait beaucoup plus de ¹²C (et ¹³C) dans l’atmosphère mais pas plus de ¹⁴C, car sa proportion est déterminée uniquement par sa production et sa désintégration.
Avec plus de CO₂ total, la même production de ¹⁴C était plus diluée et le ratio ¹⁴C/¹²C était plus faible qu’aujourd’hui. Et encore, le champ magnétique était plus fort dans le passé (on observe une perte de 5 à 10% par siècle), cela signifie qu’il y avait moins de rayons cosmiques atteignant l’atmosphère et donc encore moins de production de ¹⁴C. D’autres arguments permettent encore de supposer qu’il y avait moins de ¹⁴C dans le passé et plus de ¹²C dans le passé. Par exemple le champ magnétique était plus fort et bloquait davantage de rayons cosmiques, il y avait donc moins de production de ¹⁴C.
Le monde avant le Déluge aurait eu une végétation luxuriante (forêts immenses, climat global plus doux). Cela implique une biosphère très riche en carbone 12 (plantes, sols, etc.). Lors du Déluge, une grande partie de cette biomasse a été enfouie, devenant charbon, pétrole et gaz. Cela aurait retiré du cycle une énorme masse de carbone, modifiant ensuite le ratio ¹⁴C/¹²C.
C’est un point important, cela signifie que les échantillons anciens démarrent avec un déficit en ¹⁴C par rapport aux échantillons modernes (en réalité après 1400 av. J-C), ce qui fausse les datations (elles paraissent trop vieilles). Prenons un cas exemple:
- Un animal est mort en 2000 av. J.-C.
- Il est donc mort il y a 4025 ans
- Admettons que nous connaissons cela par la méthode historique
Je vous épargne les calculs mais si le ratio moderne (1/10¹²) reflète la réalité de l’époque 2000 av. J-C, l’âge calculé par la méthode radiocarbone donne environ 4025 ans, c’est donc correct et la méthode mathématique rejoint la méthode historique.
Cependant, si le ratio initial dans l’atmosphère était en réalité plus faible (1/5×10¹²), pour les raisons évoquées plus haut (et plus dans l’article proposé plus bas), mais qu’on utilise à tort le ratio moderne alors l’âge apparent est d’environ 17 300 ans. Nous constatons un âge gonflé à tort.
Ainsi, les calculs radiométriques partent d’hypothèses, souvent invérifiables, elles sont logiques pour certains et contestables pour d’autres. La particularité avec le ¹⁴C (en dehors du fait qu’il fonctionne bien jusqu’à 1400 av. J-C), c’est qu’il est limité à dater des objets de moins de 60 000 ans (plus détectable par la suite). Une date obtenue de 30 000 ans n’est peut-être pas correcte comme nous l’avons vu, mais on sait qu’on est en dessous de 60 000 ans. On ne peut pas dater avec précision, mais on sait dans quelle fourchette on est.
C’est pourquoi il est significatif pour les créationnistes de retrouver une quantité substantielle de carbone 14 dans les fossiles de dinosaures8, le pétrole, le charbon et les diamants. Cela permet d’apporter une preuve de la chronologie biblique qui est dans l’ordre de grandeur des milliers d’années.
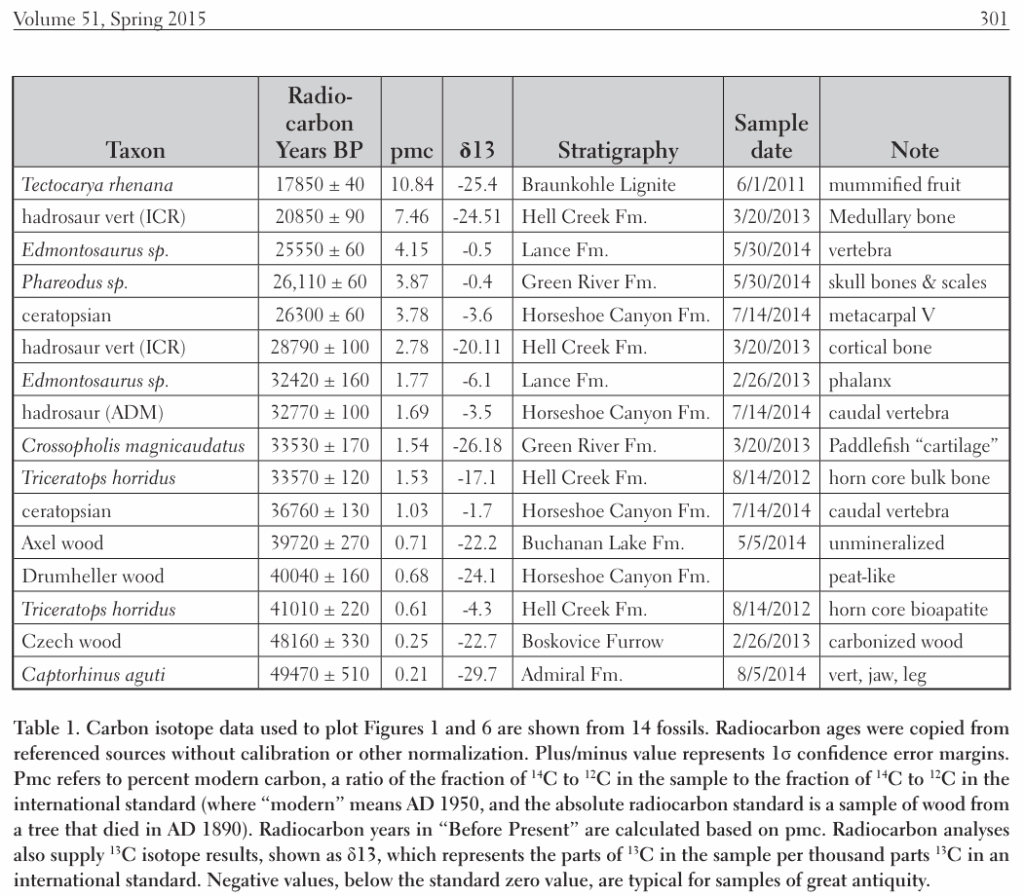
Le tableau ci-dessus de Thomas et Nelson montre un échantillon de fossiles anciens (plusieurs dizaines et centaines de millions d’années selon les évolutionnistes) avec des quantités de carbone substantielles retrouvées. Les « Radiocarbon Years BP » (before present) ne sont pas calibrées mais pourraient très bien redescendre dans l’échelle biblique en tenant compte des ratios de carbone anciens.
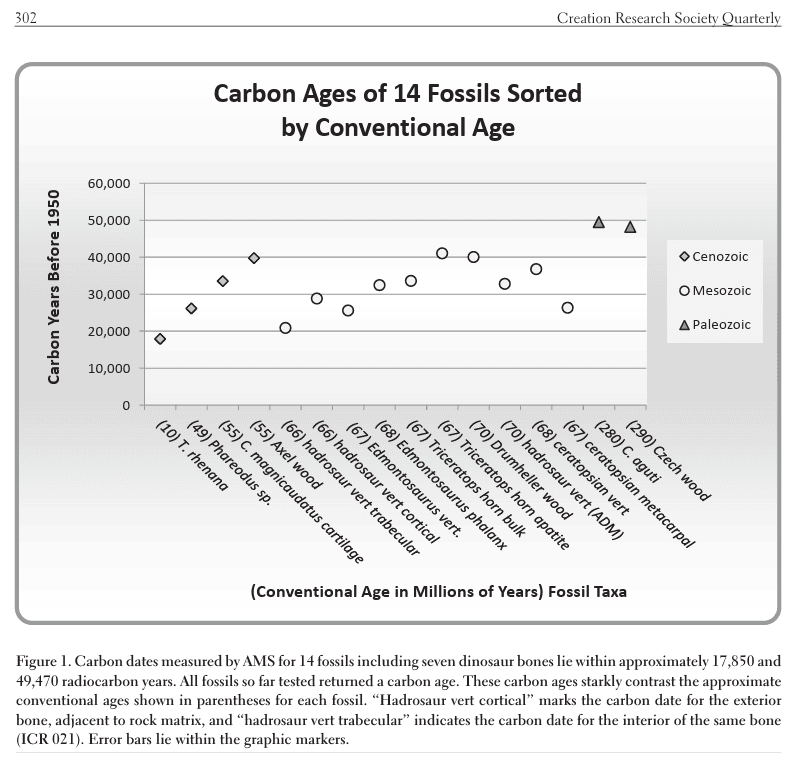
Dans l’étude, on peut lire les procédures drastiques et minutieuses utilisées pour écarter tout contaminants et ne mesurer que le carbone intrinsèque au fossile:
« Les protocoles de préparation pour l’analyse des isotopes du radiocarbone dans l’apatite osseuse ont été effectués selon Cherkinsky (2009). D’abord, les matériaux extrinsèques ont été retirés par grattage mécanique. Ensuite, les échantillons ont été laissés à tremper toute une nuit dans de l’acide acétique 1 N.
Cette étape élimine les composés carbonés qui contaminent les échantillons par colmatage sédimentaire ou par cristallisation de carbonates après dépôt. Après rinçage et séchage, environ 2 grammes d’os sont broyés puis retraités avec de l’acide acétique 1 N, avec des évacuations périodiques, jusqu’à ce que la formation de CO₂ et d’autres gaz cesse.
Ce traitement acide ne dépasse pas 72 heures, au-delà desquelles la bioapatite d’origine commence à se dissoudre, et pas seulement les matériaux carbonés secondaires de surface.
Après un nouveau séchage, plusieurs centaines de milligrammes d’os partiellement traité sont ajoutés à de l’HCl 1 N pendant moins de 20 minutes, et le CO₂ issu de la réaction est collecté. Si la masse de carbone capturé dépasse les quantités attendues, on soupçonne des apports contaminants et des traitements acides supplémentaires s’ensuivent.
Enfin, le dioxyde de carbone purifié est converti catalytiquement en graphite pour l’analyse, par spectrométrie de masse par accélérateur (AMS), du rapport 13C/14C, qui est immédiatement comparé au rapport 13C/14C de l’échantillon standard absolu de radiocarbone OX-I (NBS 4990). »
Voici deux autres citations parlantes de l’étude:
« [Andrew]Snelling a ensuite signalé un autre échantillon de bois daté au radiocarbone à 41 260 ± 540 ans radiocarbone. Trouvé près de Cripple Creek, au Colorado, il a été décrit comme étant enchâssé dans une lave datée conventionnellement par la méthode Ar–Ar à 32 millions d’années. Il a écrit : “On ne peut donc que conclure que le radiocarbone mesuré par le laboratoire doit être un radiocarbone réel, in situ, intrinsèque au bois d’origine, et non une contamination d’aucune sorte” (Snelling, 2008a). »
Il y a 4 arguments clés contre l’idée de la contamination.
- Les procédures (prétraitements acides, calibrations régulières de l’AMS) excluent rigoureusement les contaminants ; une erreur de manipulation ne pourrait pas ajouter assez de 14C moderne pour expliquer les mesures.
- Le prétraitement acide–base–acide (ABA) enlève efficacement les contaminations superficielles ; c’est la base éprouvée de la datation au 14C des os archéologiques.
- Même si des fluides hydrothermaux avaient apporté du 14C moderne, la déminéralisation rigoureuse en préparation d’échantillon au labo élimine ces apports.
- Dans le cas de Snelling, l’échantillon provenait de plus de 900 m de profondeur, rendant improbable le mélange d’eaux suffisant pour introduire autant de 14C sur des millions d’années ; en plus, le carbone dissous (soluble) des eaux n’interagit pas avec le carbone insoluble du fossile.
« Si la percolation avait contaminé ces fossiles, leurs parties externes devraient montrer des âges au radiocarbone plus faibles que leurs parties internes, mais nous n’avons pas constaté cela. »
« Nous avons envisagé la possibilité qu’une erreur puisse provenir d’un laboratoire de datation au radiocarbone donné, par exemple une erreur d’opérateur un jour donné.
Nos 16 résultats, issus d’un laboratoire d’isotopes du carbone spécialisé dans le tissu osseux, s’ajoutent à des détections similaires de radiocarbone dans des fossiles du Phanérozoïque réalisées par Geochron Laboratories (Massachusetts), l’IsoTrace Radiocarbon Laboratory de l’Université de Toronto (Ontario), l’Australia Nuclear Science and Technology Organisation (ANSTO) et le système AMS d’Iéna du Max-Planck-Institut für Biogeochemie.
L’explication la plus parcimonieuse du fait que cinq laboratoires différents détectent du radiocarbone dans des fossiles anciens semble être que ces fossiles contiennent du 14C endogène. »
Plus de détails dans les articles suivants:
- La Datation au Carbone 14 : Une Approche Créationniste (argument évolutionniste de la contamination traité)
- Le Carbone 14 dans les Diamants, le Charbon, les Fossiles et les Dinosaures
Les biomatériaux
Un autre argument soutenant indépendamment l’argumentaire sur le carbone 14 est celui des biomatériaux retrouvés dans les fossiles anciens et notamment ceux de dinosaures.9
Des restes de biomatériaux mous (vaisseaux sanguins, protéines, collagène, kératine, ADN fragmentaire, pigments comme la mélanine) ont été retrouvés dans des fossiles:
- Dinosaures (os de T. rex, hadrosaures, etc.)
- Poissons
- Amphibiens
- Oiseaux fossiles
- Invertébrés marins (trilobites, ammonites, etc.)
Ces découvertes ont été faites sur tous les continents et dans pratiquement toutes les étapes géologiques (Cambrien, Jurassique, Crétacé, Tertiaire, etc.). Comme avec le carbone 14, ces biomatériaux ne peuvent pas subsister plus de quelques milliers ou dizaines de milliers d’années.
La dégradation des biomolécules est bien documentée, et même dans des environnements optimaux, elles ne devraient pas durer plus de quelques dizaines de milliers d’années au maximum. La présence de tissus mous et de protéines renforce l’idée que ces fossiles ont été emportés il y a quelques milliers d’années lors du déluge de Noé.
« La récupération de tissus mous et de cellules à partir d’os fossiles devient de plus en plus courante, avec des structures morphologiquement compatibles avec des ostéocytes de vertébrés, des vaisseaux sanguins, une matrice fibreuse/collagénique, et de potentiels contenus intravasculaires, désormais reconnus dans des spécimens remontant jusqu’au Permien. »
— Ullmann et al., Cretaceous Research, vol. 99, 2019
Ces découvertes ont provoqué une « consternation immense » dans la communauté scientifique. Lorsque Mary Schweitzer (première découvreuse en 1990 puis dans les années 2000) a rapporté qu’elle avait trouvé des ostéocytes (et non seulement leurs empreintes fossilisées), Jack Horner était sceptique. Il pensait qu’elle faisait référence aux lacunes (les espaces vides laissés par les cellules disparues). Schweitzer a confirmé qu’il s’agissait bien de cellules intactes, avec des structures internes préservées, comme des filopodes et des noyaux.
Le superviseur de Schweitzer, Jack Horner, a écrit :
« Quand Mary travaillait pour la première fois sur ce matériau, elle m’a appelé pour me dire qu’elle avait trouvé des ostéocytes. J’ai supposé qu’elle voulait parler des espaces où les ostéocytes auraient été, ce que j’ai suggéré. »
« Non, Jack, en réalité, nous avons les cellules, et elles ont des filopodes, et elles ont des noyaux. »
« Mary, les fichus créationnistes vont juste t’adorer. »
« Jack, c’est ton dinosaure. »
Horner, J. and Gorman, J., How to build a dinosaur: The new science of reverse evolution, pp. 80-81, Dutton, 2009.
Cela illustre à quel point la découverte scientifique était inattendue dans un cadre évolutif et comment elle se corrélait naturellement au créationnisme. Alors que les évolutionnistes cherchent des explications naturelles à ce phénomène, les créationnistes revendiquent une preuve de la chronologie biblique de moins de 10 000 ans.
La progression mer-terre
L’idée que l’ordre du registre fossile reflète l’ordre dans lequel différents écosystèmes ont été enterrés pendant le Déluge est connue sous le nom de théorie de la zonation écologique. Le créationniste Harold Willard Clark l’a proposée pour la première fois en 1946 dans un livre intitulé « The New Diluvialism ». D’autres créationnistes l’ont depuis développée et modifiée.
Nous savons aujourd’hui que la Terre a une structure en couches constituée d’un noyau, d’un manteau et d’une croûte. La croûte est la fine couche extérieure et est fragmentée en une série de plaques rigides capables de se déplacer les unes par rapport aux autres.
Les découvertes de la géologie moderne, combinées aux indices de la Bible, suggèrent que ces plaques tectoniques ont été fracturées à l’époque du Déluge de Noé. Pendant le Déluge, trois événements se sont produits simultanément:
- La croûte océanique d’avant le Déluge s’est détachée des marges du supercontinent et a commencé à plonger dans le manteau terrestre, écartant les plaques et provoquant la dislocation de la masse terrestre d’avant le Déluge.
- Le plancher océanique plongeant s’est enfoncé rapidement dans le manteau, repoussant le matériau chaud environnant et agitant le manteau dans toute sa profondeur.
- Le matériau chaud du manteau est remonté (comme un tapis roulant) pour former une nouvelle croûte océanique le long des dorsales médio-océaniques où les plaques se séparaient.
Puisque la nouvelle croûte océanique était plus chaude (et donc moins dense) que l’ancienne croûte océanique froide qu’elle remplaçait, elle était plus flottante et se trouvait plus haut dans le manteau terrestre. Par conséquent, le niveau du plancher océanique a considérablement augmenté, rendant les bassins océaniques moins profonds et déplaçant l’eau vers les continents.
Le niveau de la mer a pu monter de plus d’un kilomètre (1,6 kilomètre) par rapport à son niveau d’avant le Déluge, suffisamment pour inonder les continents et recouvrir les sommets des plus hautes montagnes antédiluviennes.
De plus, le long des dorsales médio-océaniques où le nouveau matériau chaud entrait en contact avec l’eau froide de l’océan, l’eau était vaporisée et projetée dans l’atmosphère. En conséquence, des jets de vapeur supersoniques ont éclaté le long de milliers de kilomètres de dorsale médio-océanique. Ces geysers étaient probablement parmi les sources du grand abîme mentionnées dans Genèse 7:11.
L’eau océanique captée par ces jets supersoniques retombait sur la Terre sous forme d’une pluie intense et globale. Cela pourrait avoir été la principale source de la pluie qui est tombée des « écluses des cieux » pendant quarante jours et quarante nuits (Genèse 7:4,11).
Et pendant tout ce temps, alors que le monde était sous l’eau, des mouvements tectoniques catastrophiques réarrangeaient rapidement les fragments brisés du supercontinent, les séparant et les heurtant à nouveau avant de finalement les séparer pour produire les continents actuels.
À mesure que les eaux du déluge s’avançaient sur le supercontinent d’avant le Déluge, des communautés d’organismes étaient emportées, transportées et enterrées dans l’ordre dans lequel elles étaient emportées par le Déluge. De cette manière, une série horizontale de biomes d’avant le Déluge a été enterrée pendant le Déluge, produisant une séquence verticale de couches rocheuses avec des fossiles caractéristiques.
Les premiers à être submergés furent les récifs de stromatolites qui formaient une barrière autour de parties de Rodinia, séparant l’océan profond du plateau marin peu profond. Cette barrière a été franchie lorsque les marges continentales se sont effondrées au début du déluge. Ensuite, les eaux montantes ont envahi, submergeant les animaux vivant dans la lagune marine derrière le récif : d’abord les Édiacariens, puis les petites créatures à coquilles, et enfin les animaux atdabanien. L’enterrement de la faune atdabanienne du Cambrien inférieur explique l’apparition soudaine dans le registre fossile d’animaux à coquilles dures abondants et diversifiés, un événement que les géologues appellent l’explosion cambrienne.
L’ordre d’enfouissement des biomes est à peu près le suivant:
- Biomes marins profonds et côtiers (Cambrien–Dévonien)
- Biomes marins peu profonds et zones marécageuses (Carbonifère–Permien)
- Biomes continentaux intérieurs – plaines et forêts (Mésozoïque : Trias–Crétacé)
- Biomes de mammifères et d’humains (Cénozoïque : Tertiaire–Quaternaire)
Le schéma simplifié de l’ordre d’enfouissement des groupes d’organismes est le suivant:
- Océans profonds: invertébrés marins (trilobites, coraux, éponges).
- Plateformes continentales / marécages: forêts de fougères, charbon, amphibiens, reptiles.
- Plaines continentales: dinosaures, plantes terrestres, oiseaux.
- Régions plus élevées: mammifères, oiseaux, humains (dernier stade).
Par exemple des trilobites ont été retrouvés dans de la cendre, ils ont été soudainement fossilisés lors de coulées pyroclastiques sous-marines. Ils sont dénommés « les trilobites Pompéi ». Cela correspond probablement à la phase initiale du déluge dans les océans.10
Certains créationnistes défendent une fin du Déluge à K-Pg (crétacé/paléogène). Ils doivent alors expliquer l’apparition rapide des mammifères marins dans le Cénozoïque. Si la limite du Déluge était à K-Pg, alors tous les fossiles de baleines cénozoïques seraient des vestiges de petites catastrophes post-Déluge mais leur distribution globale et massive indique plutôt un phénomène catastrophique mondial. Cela suggère que le Déluge était toujours en cours pendant le Cénozoïque.
Beaucoup de dinosaures ont été rapidement ensevelis par des coulées de boue, tsunamis et dépôts côtiers. Leurs corps lourds, moins adaptés à flotter longtemps, ont coulé ou se sont déposés plus rapidement.
Les carcasses de cétacés ont une forte flottabilité (graisse, air emprisonné dans les poumons, structure du corps). Elles peuvent rester longtemps à la surface avant de se remplir d’eau et couler. Le résultat est qu’elles auraient été transportées plus loin et ensevelies plus tardivement dans les séquences du Déluge, souvent dans des bassins profonds.
On sait que les carcasses de baleines modernes (les fameux whale falls) peuvent flotter longtemps avant de couler. Cela cadre bien avec l’idée qu’elles auraient été enterrées après coup, donc trouvées stratigraphiquement au-dessus des dinosaures.
La disparition des dinosaures
« Une nouvelle étude montre que les dinosaures – et d’autres vertébrés continentaux – sont restés diversifiés en Europe jusqu’à l’impact de l’astéroïde, il y a 66 millions d’années. C’est une preuve solide que les dinosaures et nombre de leurs contemporains se sont éteints rapidement et simultanément partout dans le monde. »
– ScienceDaily, 14 janvier 2015
On ne croit pas en tant que créationniste à l’explication évolutionniste de l’astéroïde, mais cela montre que les séculiers ont un scénario cherchant à décrire la même chose que les créationnistes. Le Déluge de Noé est un bien meilleur candidat, il permet d’expliquer la fossilisation à l’échelle mondiale dans les diverses étapes géologiques, ce que ne peut expliquer un astéroïde qui aurait eu un effet d’incendie, de refroidissement et de bouleversement écologique n’impliquant pas nécessairement l’eau et les sédiments nécessaires à la fossilisation.
En fait, le modèle diluvialiste répond aux nombreuses interrogations évolutionnistes sur la disparition des dinosaures. Bien que la théorie de l’astéroïde soit la plus connue, une centaine d’hypothèses a été proposée pour essayer d’expliquer la disparition à l’échelle mondiale:
« Au fil des années, plus d’une centaine d’hypothèses ont été proposées pour expliquer l’extinction des dinosaures. »
Benton, M. 2015. Vertebrate Paleontology, 4ᵉ éd., p. 262.
Tous les scénarios d’extinction possibles ont été considérés (… sauf un…).
« Les preuves cumulées [d’enfouissement fossile] indiquent que les individus d’Edmontosaurus sont morts lors d’un événement de mortalité de masse et que leurs restes désarticulés ont été rapidement ensevelis dans un étang peu profond de plaine d’inondation… »
— Ullmann et al., Cretaceous Research, vol. 99, 2019
L’Alberta (Canada) dévoile une « momie de dinosaure » retrouvée avec la peau et le contenu intestinal intacts:
« Âgé de “110 millions d’années”, pris dans une rivière et emporté vers la mer »
— Greshko, M., 2011, National Geographic.
« Beaucoup pensent que de vastes inondations ont emporté les dinosaures, causant leur mort. »
— Canadian Geographic, 2014
« Les restes fossiles [de dinosaures momifiés] sont d’un réalisme saisissant, ressemblant à un dragon endormi. Selon National Geographic, qui a financé la préparation du fossile pendant cinq ans (7 000 heures), il est probable que la créature de 3 000 livres et 18 pieds de long (environ 1 360 kg et 5,5 m) soit morte dans ou près d’une rivière. Sa carcasse gonflée a ensuite dérivé vers la mer avant de couler, dos en premier, dans la vase où la fossilisation a commencé. »
Daley, J. Spectacularly Detailed Armored Dinosaur “Mummy” Makes Its Debut. Smithsonian. Publié sur smithsonianmag.org 15 Mai, 2017
« C’est difficile à imaginer. Cet animal [un Hadrosaure] est mort il y a 76 millions d’années. Il a été parfaitement conservé depuis, et il se trouve qu’au moment où nous passions, il commençait tout juste à s’éroder hors de cette falaise… Cet animal est probablement soit mort puis immédiatement recouvert de sable et de limon dans la rivière, soit tué parce qu’une berge s’est effondrée sur lui. »
Mayorquin, O. ‘Dinosaur mummy’: Researchers believe they found one of the best preserved dinosaurs ever. USA Today. Publié sur usatoday.com 14 septembre, 2022
Frank Sherwin déclare:
« On se rappelle qu’un déluge d’ampleur épique — et non une rivière ou sa berge — pourrait submerger rapidement la créature sous du limon et du sable, entraînant une conservation exceptionnelle. En effet, au fil des décennies, d’importants progrès de la théorie de la sédimentation ont conduit les géologues à reconnaître que la plupart des unités rocheuses résultent de processus étendus et de forte intensité. Des phénomènes géologiques traditionnellement attribués à des actions graduelles et lentes se sont révélés pouvoir s’accomplir en quelques minutes à peine. Tout cela pointe vers le Déluge rapporté dans la Genèse. »
Joel Tay donne 7 preuves que les ossements de dinosaures sont une preuve du déluge mondial:
- Les dépôts massifs et rapides de milliers d’ossements de dinosaures
- La pose opisthotonique (pose de la mort qui indique qu’ils sont morts dans l’eau, tête arquée vers l’arrière, et queue vers l’avant due à une contraction)
- Les dinosaures à plaques (blindés) fossilisés à l’envers, c’est à dire ventre vers le haut et dos vers le bas (preuve qu’ils sont morts dans l’eau, la lourdeur du blindage sur leur partie haute fait qu’ils se sont retournés, par exemple lorsqu’une vague les a fait basculer sur leur dos)
- Les dinosaures fossilisés en plein combat (ou chasse) ce qui montre qu’ils ont été fossilisés en un éclair
- Les couches polystrates d’œufs de dinosaures (plusieurs œufs disposés en cercle sur une hauteur d’une quinzaine de cm ce qui montre la rapidité de la sédimentation, la maman dinosaure pondait alors que les sédiments s’accumulaient rapidement, aucun temps d’exposition qui aurait détruit les œufs)
- Le carbone 14 dans les fossiles de dinosaures (d’autant plus significatif que les dinosaures en haut de la colonne géologique ont les mêmes quantités détectées que les dinosaures qui sont plus bas, entre 20 000 et 50 000 ans non recalibrées, ce qui appuie l’idée d’un même évènement commun qui les a fossilisés)
- Les tissus mous et biomatériaux dans les fossiles de dinosaures
Légendes de déluge
Il existe plus de 300 récits traditionnels de déluge collectés dans les cultures du monde entier. Ces récits apparaissent sur tous les continents: Mésopotamie, Afrique, Amérique du Nord et du Sud, Asie, Océanie, Europe. Cela appuie que l’humanité entière a gardé une mémoire commune du cataclysme mondial.
La plupart des récits de déluge contiennent des éléments parallèles à Genèse 6–9:
- Un jugement divin ou colère des dieux.
- Un homme juste/choisi averti de construire un bateau.
- Sauvetage de la famille du héros.
- Sauvetage d’animaux pour repeupler la Terre.
- Une embarcation qui flotte sur les eaux.
- L’eau couvre toute la terre (souvent décrit comme universel).
- Après le retrait des eaux, l’homme offre un sacrifice.
- Une nouvelle humanité issue des survivants.
Les ressemblances ne semblent pas être une « coïncidence culturelle », mais dues à des variations régionales d’un même souvenir historique commun. La divergence entre les récits s’explique par la dispersion des peuples après Babel (Genèse 11). Chaque peuple a transmis l’histoire du Déluge, avec des altérations locales (folklore). L’uniformité des thèmes (bateau, famille, animaux, jugement, sacrifice) renforce la thèse d’un événement réel universel.
Un documentaire diffusé il y a quelques années explore le sujet en détail, en particulier le lien supposé avec l’Épopée de Gilgamesh.
Pictogramme chinois
Les pictogrammes chinois semblent contenir des éléments du récit de la Genèse. L’un des caractères les plus connus sur ce sujet est celui pour « bateau ». Le caractère chinois 船 (chuan) est composé de trois éléments:
- 舟 (navire)
- 八 (huit)
- 口 (bouches et classificateur pour personne)
Cela nous rappelle que 8 personnes ont été sauvées sur l’Arche de Noé. Une diversité de caractères chinois (dont l’origine remonte à Babel selon le modèle biblique) contient des éléments de la Genèse de manière qui ne peut simplement être expliqué par la coïncidence.
Echos de la tour de Babel
Certains récits diluviens semblent même faire allusion à l’épisode de la Tour de Babel avec la dispersion des peuples et la création des langues:
Par ailleurs les textes écrits plus anciens remontent à environ 3500-4500 ans (sumérien cunéiforme) en arrière ce qui est cohérent avec le redémarrage de l’humanité après le déluge en 3300 av.J-C.
- Quand a eu lieu le déluge biblique? En quelle année?
- Les Anneaux des Arbres et la Chronologie Biblique
Les langues
La Bible enseigne que Dieu a créé les langues distinctes à Babel, il y a environ 5 000 ans (Genèse 11). Cela a forcé la dispersion des peuples et leur migration, conformément à l’ordre divin de remplir la terre. Les linguistes évolutionnistes pensent que toutes les langues actuelles ont évolué à partir de proto-langues depuis environ 12 000 ans.
Edward Vajda (Université de l’Alaska) a démontré une relation statistique forte entre deux familles linguistiques:
- Yeniseian (Sibérie, ex. langue Ket, ~200 locuteurs).
- Na-Dené (Amérique du Nord, ex. Navajo).
Jared Diamond (dans Nature) souligne que ces langues partagent une structure grammaticale trop proche pour avoir divergé il y a 12 000 ans. Normalement, après 5 000–10 000 ans, les langues deviennent méconnaissables entre elles.
Si ces langues se ressemblent encore aujourd’hui, c’est qu’elles ont divergé bien plus récemment que 12 000 ans. Cela correspond à la chronologie biblique avec la dispersion depuis Babel il y a environ 5 000 ans. Les grandes familles linguistiques (Na-Dené, Yeniseian, etc.) seraient issues directement des langues créées à Babel, et leur relative proximité confirme une origine commune récente.
Après le Déluge, lors de la période glaciaire postdiluvienne, le niveau marin était plus bas (environ 100 m de moins). Un pont terrestre au détroit de Béring permettait la migration de familles humaines d’Asie vers l’Amérique (et potentiellement d’animaux). Cela explique la diffusion des langues apparentées entre Sibérie et Amérique du Nord.
Les linguistes recensent aujourd’hui 6 000 à 7 000 langues vivantes. Celles-ci se regroupent en environ 70 grandes familles linguistiques (certains parlent de 60–90, selon les classifications). Voici quelques exemples:
- Indo-européen (français, anglais, grec, hindi, etc.)
- Sino-tibétain (chinois, tibétain, birman…)
- Afro-asiatique (hébreu, arabe, berbère…)
- Bantou/Niger-Congo (swahili, zoulou, etc.)
- Na-Dené (navajo, apache…)
- Yeniseian (Ket en Sibérie)
Les linguistes évolutionnistes postulent une proto-langue unique (souvent appelée Proto-World) mais les 70 familles principales ne peuvent pas être reliées les unes aux autres par une reconstruction linguistique fiable (un peu comme les grands groupes d’organismes dans le registre fossile). Même les comparaisons les plus poussées échouent au-delà de 5 000 à 10 000 ans.
Les familles linguistiques apparaissent comme indépendantes, chacune avec ses propres structures et racines. De plus plus on remonte loin plus les langues sont complexes (beaucoup plus de formes verbales, par exemple le grec ancien est plus complexe que le latin qui est plus complexe que les langues qui en ont dérivées).
La Bible dans Genèse 11 décrit 70 nations après Babel (voir le Tableau des Nations en Genèse 10). Des créationnistes comme Henry Morris et Bodie Hodge ont souligné que ce nombre correspond étonnamment bien aux grandes familles de langues recensées aujourd’hui.
L’explication est qu’à Babel, Dieu n’a pas « fait évoluer » une langue unique en plusieurs dialectes. Il a créé de toutes pièces des langues radicalement différentes, incompréhensibles entre elles. Chaque famille est l’héritière d’une de ces langues originelles. Retrouvez toutes ces informations à l’article suivant:
Récits de dragons
Les récits de dragons pullulent sur tous les continents avec des noms différents. Leur description « reptilienne », associé à certaines trouvailles archéologiques, permettent d’appuyer une fois de plus l’existence « historique » des dinosaures, il y a quelques milliers d’années.

On peut observer l’emblème de la « salamandre » couronnée de François Iᵉʳ (avec des flammes). On le voit notamment dans la Galerie François-Iᵉʳ au château de Fontainebleau, et aussi à Chambord et Blois.

Des « dinosaures » historiques auraient pu projeter un jet chaud/flammable par un mécanisme analogue au coléoptère bombardier avec un mélange de réactifs stockés séparément puis expulsés sous pression. Les idées incluent des gaz intestinaux (méthane) entraînant de très petites quantités de phosphine/diphosphane qui s’enflamment spontanément à l’air ; une décharge électrique comme amorce d’allumage (analogie lointaine avec l’anguille électrique) ; des cavités crâniennes (p. ex. chez certains hadrosaures) servant de « chambre de mélange ».
La ressemblance dragons/dinosaures est nettement perçue chez beaucoup de chercheurs et historiens:
« Les dragons des légendes ressemblent étrangement à de véritables créatures qui ont vécu dans le passé. Ils sont très semblables aux grands reptiles qui habitaient la Terre bien avant l’époque à laquelle on suppose que l’homme est apparu. Les dragons étaient généralement mauvais et destructeurs. Chaque pays en avait dans sa mythologie. »
— Wilson, Knox. 1973, World Book Encyclopedia, vol. 5.
Carl Sagan, un gagnant du prix Pulitzer, qui a écrit le livre : The Dragons of Eden, a même tenté d’expliquer que la récurrence des dragons à travers les différentes cultures du monde, était due à un souvenir que nous aurions gardé depuis l’époque où nous étions encore des mammifères, lorsque les dinosaures existaient, et qu’à travers notre ADN nous en aurions gardé le souvenir, sans pour autant que nous en ayons réellement vu.
Il explique que ce serait la peur horrible provoquée qui se serait imprimée dans notre cerveau à travers des millions d’années et qui subsisterait encore aujourd’hui.
Un nouveau fossile « Dracorex » constitue une forte indication que les dragons étaient réels, et que le récit biblique de dinosaures et d’humains vivant ensemble est vrai.

« La forme du crâne du dinosaure, avec son long museau, ses bosses et ses cornes bizarres, a surpris les scientifiques, » a-t-elle déclaré [Adrienne Mayor, chercheuse invitée à Stanford.]. « Mais ce crâne paraît étrangement familier à quiconque a étudié les dragons ! Dracorex présente une ressemblance frappante avec les dragons de la Chine ancienne et de l’Europe médiévale. »
Hérodote, surnommée le « père de l’histoire », a aussi parlé des dragons (deuxième livre) :
On dit que les serpent ailés volent d’Arabie au début du printemps, et vont vers l’Égypte. Ils sont comme des serpents d’eau. Ils n’ont pas de plumes sur leurs ailes mais sont plutôt comme les ailes des chauves-souris.
Les descriptions qui sont faites de ces créatures dans les différentes cultures est un témoignage puissant que les dinosaures ont vécu en même temps que toutes les autres espèces, y comprit les humains. Ces derniers ont fini par disparaître (avec quelques derniers spécimens de petite taille au moyen-âge) probablement pour des raisons de climat et de réduction de leur habitat (assèchement des zones marécageuses par exemple).
- Les Dinosaures ont-ils vécu et disparu il y a des millions ou quelques milliers d’années ? Créationnisme
- Les Dragons étaient-ils des « Dinosaures Historiques » ?
Ere glaciaire
Le déluge avec son activité volcanique intense a laissé les océans chauds et les continents froids (aérosols bloquant le rayonnement solaire). Cela a provoqué des conditions uniques et spécifiques ayant permis une évaporation massive d’eau océanique retombant sous forme de neige sur les continents selon les latitudes. Les étés étaient frais et ne permettaient pas de faire fondre la glace. Cet âge de glace aurait duré selon le météorologue créationniste, Michael Oard, de 250 à 1300 ans11.
Dans certains endroits côtiers, les températures auraient paradoxalement été plus chaudes qu’aujourd’hui car l’océan chaud réchauffait le climat. Les régions côtières baignées par des océans encore chauds auraient connu des hivers plus doux et parfois même des températures supérieures à l’actuel. Cela explique pourquoi, dans certains dépôts glaciaires, on trouve des zones tempérées voisines de zones englacées. Les océans chauds étaient la source d’humidité qui alimentait les énormes chutes de neige sur les continents refroidis.
Aujourd’hui, la Sibérie est trop froide, trop sèche, et trop pauvre en végétation pour supporter de grandes populations de mammouths mais juste après le Déluge, les océans encore chauds réchauffaient les côtes nordiques et alimentaient une humidité intense. Cela donnait des étés plus doux et humides qu’aujourd’hui, produisant assez de végétation pour nourrir des troupeaux entiers de mammouths, rhinocéros laineux, etc.
Ensuite, quand l’océan s’est refroidi et que les hivers se sont durcis, ces animaux ont été piégés dans des tempêtes soudaines, ce qui explique la congélation rapide de certains mammouths retrouvés intacts.
On a trouvé des fossiles d’hippopotames en Grande-Bretagne, en Allemagne et jusqu’en Scandinavie12. Dans le modèle classique, cela semble contradictoire: comment des animaux tropicaux pouvaient-ils vivre dans une Europe glaciaire?
Dans le modèle post-Déluge, les océans chauds rendaient les régions côtières d’Europe bien plus tempérées qu’aujourd’hui. Les hippopotames pouvaient prospérer dans ces zones de rivières et lacs relativement chauds, même pendant que l’intérieur des continents se refroidissait.
Biogéographie
L’ère glaciaire, en faisant baisser le niveau des mers, a provoqué des ponts terrestres entre les continents, permettant la migration mondiale des animaux. La Bible indique que tous les animaux terrestres ont survécu au Déluge dans l’arche de Noé, puis se sont dispersés à partir du Moyen-Orient. Cela explique pourquoi on retrouve des « points de radiation » dans les archives fossiles et actuelle: les groupes apparaissent soudainement et se diversifient. Par exemple les marsupiaux d’Australie n’ont pas toujours été là, mais y ont migré via des couloirs terrestres ou des radeaux de végétation.
Des migrations rapides ont été possibles:
- Les marsupiaux ont atteint l’Australie (via l’Asie du Sud-Est).
- Les mammouths et bisons sont passés en Amérique du Nord via le détroit de Béring.
Les populations isolées ont ensuite divergé rapidement (spéciation post-Déluge), ce qui explique la variété géographique. La biogéographie fossile montre des animaux dans des régions où ils ne pourraient pas vivre aujourd’hui:
- Hippopotames en Angleterre et en Allemagne (climat tempéré post-Déluge grâce aux océans chauds).
- Mammouths en Sibérie (été doux et humide grâce à l’océan chaud, puis refroidissement brutal).
Cela colle avec le modèle d’ère glaciaire unique post-Déluge décrit par Michael Oard.
Des îles comme les Galápagos ou Madagascar montrent une grande diversité d’espèces proches (pinsons, lémuriens, caméléons…).
Certaines répartitions complexes s’expliquent potentiellement avec le scénario terre jeune, par exemple les singes d’Afrique et d’Amérique du Sud sont séparés par l’Atlantique. Les rongeurs et paresseux ont eux aussi traversé l’océan pour coloniser l’Amérique du Sud. Les radeaux végétaux massifs laissés par le déluge sont bien plus adaptés à la tâche que les radeaux évolutionnistes produits par de simples tempêtes.
Les animaux sortant de l’Arche (au Moyen-Orient) pouvaient migrer plus facilement si les continents étaient encore semi-connectés ou proches. Certaines routes pouvaient exister sous forme de ponts terrestres temporaires (effet combiné de continents pas encore trop éloignés + niveau marin plus bas durant l’âge glaciaire).
Dans le modèle créationniste, la séparation finale des continents (rupture de la Pangée et ouverture de l’Atlantique) s’est achevée juste après le Déluge, durant l’ère glaciaire post-déluge. Cela veut dire que les organismes fossilisés avant ou pendant le Déluge pouvaient appartenir à une biosphère encore largement pangéenne (continents connectés). Cela résout par exemple l’énigme d’un primate africain retrouvé en Amazonie péruvienne13.
Le fait de retrouver par exemple des fossiles de singes ou de rongeurs “en Afrique” et d’autres “en Amérique du Sud” ne reflète pas une séparation biogéographique réelle à leur époque vivante. Ils pouvaient très bien cohabiter sur un supercontinent avant que l’Atlantique ne s’ouvre.
Pour les créationnistes, la grande majorité des fossiles (du Cambrien au Pliocène) ont été déposés pendant le Déluge. Ces organismes ont été enterrés dans des sédiments catastrophiques, déplacés par les eaux, la tectonique rapide et les courants. Leur position géographique dans les strates ne reflète pas forcément l’endroit où ils vivaient. Par exemple un dinosaure trouvé dans le Montana ne signifie pas que cet animal vivait exactement à cet endroit-là, mais qu’il a été transporté, enseveli et fossilisé là durant les dépôts cataclysmiques.
Les fossiles et les espèces actuelles montrent que les marsupiaux ne se limitent pas à l’Australie. L’opossum par exemple vit encore aujourd’hui en Amérique. Dans le registre fossile, on retrouve des marsupiaux en Amérique du Sud et même en Europe. Dans le modèle créationniste, cela s’explique par une dispersion post-Déluge à partir du Moyen-Orient, les marsupiaux auraient migré vers l’Australie en suivant des corridors terrestres (via l’Asie du Sud-Est), mais certains groupes se seraient arrêtés ou maintenus en Amérique du Sud.
Il existe des peintures rupestres en Inde (district de Hampi et Madhya Pradesh) représentant des animaux ressemblant fortement à des kangourous14. Or, les kangourous sont aujourd’hui endémiques à l’Australie. Cela s’interprète directement dans un scénario post-Déluge, les marsupiaux auraient migré à travers l’Asie du Sud (l’Inde est sur la route menant de la Turquie à l’Australie), et certaines populations (aujourd’hui disparues) auraient laissé une trace en Inde avant de disparaître, tandis que la branche principale atteignait l’Australie via des ponts terrestres (qui existaient à l’époque de l’ère glaciaire).
L’ocre (surtout rouge, à base d’oxydes de fer) est utilisée presque partout où l’on trouve de l’art rupestre et des pratiques symboliques anciennes. On en a des preuves fortes en Afrique (Blombos, Pinnacle Point), en Asie (Sulawesi), en Europe, en Australie (Arnhem Land, sépultures de Lake Mungo) et dans les Amériques (Cueva de las Manos, Argentine).
Dans un cadre créationniste, on peut proposer qu’un « kit culturel » commun (dont l’ocre) était connu et utilisé dans l’humanité unifiée avant la dispersion de Babel. Cette technologie s’est diffusée avec les populations post-Babel, ce qui cohérent avec l’omniprésence du pigment. Par la suite, l’ocre étant facile à découvrir, la technique a continué d’être utilisée pendant des siècles.
- Réponse créationniste à la vidéo « les preuves de l’évolution » de Passe-Science – sur la répartition géographique (7)
- L’Homo erectus est-il une transition entre l’homme et un singe?
Perte de technologies (chasseurs-cueilleurs, agriculteurs)
L’étude de Julio Mercader (2009), publiée dans Science15, a montré que des Homo sapiens du « Middle Stone Age » au Mozambique (datés dans le cadre évolutif à environ 105 000 ans) consommaient déjà régulièrement des graines de sorgho sauvage et d’autres végétaux (palmiers, bananiers, tubercules).
70 outils en pierre (environ 12 % de l’ensemble retrouvé) ont été sélectionnés pour l’analyse, incluant des grattoirs, outils à moudre, pointes, éclats et autres. Les analyses d’outils en pierre ont révélé des milliers de granules d’amidon, dont une part significative montrait des traces de broyage ou de cuisson, preuve d’une préparation culinaire sophistiquée. Cette découverte repousse très loin dans le passé l’usage de céréales, bien avant les 10 000 ans classiquement attribués à la naissance de l’agriculture.
Du point de vue biblique, cela s’accorde avec l’idée que l’homme, dès sa création, possédait l’intelligence et les compétences techniques pour transformer et utiliser une large variété de ressources alimentaires, conformément au mandat donné en Genèse de dominer la terre et de se nourrir des plantes. Cela montre que la maîtrise des céréales et des outils n’est pas l’aboutissement lent d’une évolution culturelle, mais une capacité originelle de l’humanité.
L’anthropologue Julio Mercader a découvert qu’« un vaste assemblage de granules d’amidon a été retrouvé à la surface d’outils en pierre de l’âge de pierre moyen au Mozambique, montrant que les premiers Homo sapiens se nourrissaient de graines de graminées dès il y a au moins 105 000 ans, y compris celles des sorghos. »16
Le scénario dominant place la « révolution néolithique » (naissance de l’agriculture) vers 10 000 ans avant aujourd’hui, avec les premières cultures de céréales au Proche-Orient (blé, orge) mais si des humains utilisaient déjà des graines de sorgho il y a 100 000 ans, cela repousse très loin l’idée que l’homme ne savait rien des céréales avant le Néolithique. Cela met à mal l’idée d’une rupture nette entre « chasseurs-cueilleurs et agriculteurs » et suggère plutôt une continuité dans la connaissance et l’usage des plantes.
Il n’y a pas de raisons de penser que l’homme ait eu besoin de beaucoup de temps pour comprendre que les plantes poussaient à partir des graines. Dans Genèse, Adam reçoit immédiatement la tâche de s’occuper du Jardin d’Eden, puis il est question après le péché que c’était à la sueur de son visage qu’il mangerait du pain (Gen 3:19).
Les Mlabris (ou Phi Tong Luang), une petite ethnie vivant en Thaïlande et au Laos, ont longtemps été décrits comme « primitifs », sans agriculture ni habitat durable, vivant de cueillette et de chasse dans la forêt.
Les Mlabris ne sont pas un « peuple fossile », mais plutôt un groupe issu de populations agricoles qui ont perdu leurs techniques de culture. Leur pauvreté en outils et en organisation n’est pas une survivance préhistorique, mais une dégringolade culturelle due à l’isolement, à la marginalisation et à la perte de savoirs transmis.
Des recherches génétiques (par ex. Oota et al., 2005, Science) ont confirmé que les Mlabris sont très proches de populations agricoles voisines, et qu’ils se sont séparés récemment (peut-être seulement depuis 500 à 1000 ans).
Après Babel, l’humanité a été dispersée « selon leurs langues » (Genèse 11). Certains groupes se sont retrouvés isolés, avec des ressources limitées. Certains ont conservé un haut niveau de technologie (civilisations antiques en Égypte, Mésopotamie, etc.), tandis que d’autres ont perdu des savoirs (agriculture, métallurgie, écriture).
Au lieu d’une lente progression du « primitif vers le civilisé », le modèle créationniste décrit une perte de connaissances après la dispersion. Le cas des Mlabris montre comment une population agricole a pu devenir « chasseuse-cueilleuse » après une perte de technologies: cela reflète un scénario qui a pu se produire après la dispersion de Babel.
Défenses évolutionnistes
- Réponse créationniste à la vidéo « les preuves de l’évolution » de Passe-Science – sur les preuves paléontologiques (4)
- L’Évolution et l’épique quête des fossiles intermédiaires et des ancêtres évolutionnaires
- Réponse créationniste à la vidéo « les preuves de l’évolution » de Passe-Science – sur la répartition géographique (7)
- L’Australopithèque est-il l’ancêtre de l’homme?
- https://en.wikipedia.org/wiki/Skeletonization.
- Syme, C.E. and Salisbury, S.W., Patterns of aquatic decay and disarticulation in juvenile Indo-Pacific crocodiles (Crocodylus porosus), and implications for the taphonomic interpretation of fossil crocodyliform material, Palaeogeography, Palaeoclimatology, Palaeoecology 412:108–123, 2014.
- Allison (1988) – Décès et minéralisation en conditions anaérobies.
- Expériences avec microbial mats (Iniesto et al., 2016).
- Taphonomie expérimentale – Briggs (2016).
- Zangerl, R. and Richardson, E.S., The paleoecological history of two Pennsylvanian black shales, Fieldiana: Geology Memoirs 4, 1963.
- Gould, S.J. and Eldredge, N., Punctuated equilibrium comes of age. Nature 366:223–224, 1993.
- Thomas, B. and V. Nelson. 2015. Radiocarbon in Dinosaur and Other Fossils. Creation Research Society Quarterly. 51(4): 299-311.
- https://docs.google.com/spreadsheets/d/1eXtKzjWP2B1FMDVrsJ_992ITFK8H3LXfPFNM1ll-Yiw/edit?gid=0#gid=0.
- https://www.icr.org/content/trilobite-troubles-24.
- https://www.icr.org/content/ice-age-and-genesis-flood.
- https://creation.com/en/articles/hippos-england.
- Seiffert, E.R., Tejedor, M.F., Fleagle, J.G., Novo, N.M., Cornejo, F.M., Bond, M., de Vries, D., and Campbell Jr., K.E., A parapithecid stem anthropoid of African origin in the Paleogene of South America, Science 368:194–197, 2020.
- https://amp.scroll.in/magazine/921926/did-kangaroos-ever-live-in-india-a-new-discovery-has-archaeologists-hopping-with-excitement.
- Willyard, C. Pass the Sorghum, Caveman. Science Now. Publié sur sciencemag.org le 17 décembre 2009, accès le 4 mars 2010, concernant une étude publiée dans Mercader, J. 2009. Mozambican Grass Seed Consumption During the Middle Stone Age. Science. 326 (5960): 1680-1683.
- Mercader, J. Mozambican Grass Seed Consumption During the Middle Stone Age. Science. 326 (5960): 1680.

Inscrivez-vous sur QQLV!
Pour soutenir l’effort du ministère et la création de contenus:
RECEVEZ DU CONTENU par email
Recevez du contenu biblique, archéologique et scientifique dans votre boîte mail!